Опыт анализа причин, определяющих распространение природных очагов чумы во внетропической Евразии (В. В. Кучерук)
В наше время бурно развивается один из новых разделов географической науки - медицинская география. Она изучает географические закономерности распределения болезней человека и причины, обусловливающие приуроченность их к определенным участкам земной поверхности (Павловский, 1954).
Каждому возбудителю болезни, так же как и другим организмам свойственна определенная область распространения - ареал. Размеры и форма ареала определяются двумя основными группами причин: современными экологическими условиями, дающими виду возможность существовать на определенном участке земной поверхности, и историческими причинами, позволившими или не позволившими ему заселить подходящую по экологическим условиям территорию.
Человек оказывает на ареалы возбудителей заразных болезней несравнимо большее влияние чем на ареалы диких и даже культурных растений и животных. В отношении возбудителей трансмиссивных болезней этот вопрос в общих чертах был рассмотрен В. Н. Беклемишевым (1959). В настоящем сообщении мы ограничимся рассмотрением факторов (в том числе и деятельности человека), определяющих границы ареала возбудителя чумы только в пределах внетропической Евразии.
Воздействие человека на формирование ареалов возбудителей болезней животных, которыми он заражается и по отношению к которым может стать эпидемически значимым источником инфекции (чума, желтая лихорадка и др.), довольно велико, хотя всесветное распространение человеком возбудителей этих болезней не осуществилось. Перенос этих возбудителей с материка на материк и укоренение в новых условиях чрезвычайно затруднены. Все указания на примеры подобного расширения ареалов недостаточно научно обоснованы и носят сугубо гипотетический характер. Однако весьма вероятно, что в пределах отдельных материков возбудители этих болезней с помощью человека заняли все подходящие для их существования территории.
Чума относится к типичным природноочаговым зоонозам. Ее возбудитель постоянно циркулирует в популяциях диких грызунов некоторых ландшафтных зон. Первичное (первое в очаге) заражение человека является следствием перехода на него заразного начала от больных грызунов. Однако для чумной инфекции характерны некоторые специфические особенности, резко отличающие ее от большинства других природноочаговых зоонозов. Одной из таких особенностей является то, что, попав в человеческий коллектив, инфекция, при определенных условиях, может далее распространяться среди людей, как с помощью переносчиков - человеческих блох, а возможно и вшей (Хорхе - Jorge 1933; Блан и Балтазар - Blanc, Baltazard, 1943: Новикова и Лалазаров, 1944; Кучерук, 1949; Федоров, Рогозин, Фенюк, 1955), так и непосредственно от человека к человеку (легочная чума). Однако, несмотря на широкое распространение чумных эпидемий в недалеком прошлом, такой путь циркуляции возбудителя чумы 'является случайным, не имеющим значения для его существования как биологического вида (Жданов, 1953). Вторая особенность чумной инфекции - это ее чрезвычайно легкий переход к домовым грызунам и их блохам. Весьма важно, что даже в чуждых ландшафтных условиях эпизоотии чумы у грызунов-сожителей человека, могут тянуться довольно долгий срок - до 10 и более лет (Абе, 1942).
Согласно литературным данным Ф. Дербека (1905), Ву-Лиен-те (Wu-Lien-Teh, 1926), Р. Поллицера (Pollitzer, 1954) и др., за исторически сравнительно короткий период времени, с начала нашего летоисчисления, эпидемии чумы неоднократно покрывали почти всю территорию Евразии. За исключением северной окраины материка, трудно найти участок его, где на протяжении этого отрезка времени не было бы зарегистрировано наличие чумных эпидемий. Следовательно, возбудитель чумы при посредстве людей неоднократно распространялся по всей Евразии. Если больной или умерший человек мог служить источником возникновения эпизоотии среди местных грызунов, то возбудитель чумы имел шансы укорениться в любом участке территории, где экологические условия обеспечивали возможность его существования.
В связи с этим, следует подробнее рассмотреть вопрос, насколько велика вероятность заражения грызунов от больного человека или его трупа. Заражение зверьков может осуществиться несколькими путями: алиментарным, при рытье зараженной почзы (Балтазар, 1964) или с помощью переносчиков. Хорошо известно, что многие виды грызуноЕ - крысы, хомяки, полевки, песчанки, суслики в некоторые сезоны испытывают большую потребность в белковых кормах и охотно поедают свежие и замерзшие трупы млекопитающих. В литературе описаны прямые наблюдения за грызуном (видимо джунгарским хомячком), обгрызавшим труп умершего от чумы человека (Дудченко, 1909).
Возможность алиментарного заражения зверьков при каниба-лизме или поедании трупов, павших от чумы особей другого вида, доказана для серых крыс, мышей-малюток, когтистых, полуденных и ливийских песчанок, полевок - серебристой и Брандта (Тихомирова, 1934; Скородумов, 1936; Туманский, 1948; Тарасов, Абрамова и Демидова, 1949; Балабакин и Солодкая, 1956; Щекунова, 1957). Полевые наблюдения и специально поставленные эксперименты привели китайских ученых (Цен-Ко-сюн, 1958) к выводу, что при зимних эпизоотиях чумы среди когтистых песчанок алиментарный путь передачи инфекции является одним из основных.
По мнению В. Л. Лобанова (1956), смерть человека от чумы почти всегда есть результат гематогенной диссеминации, дающей картину быстро протекающего сепсиса. Интенсивность бактериемии при чумном сепсисе достаточна высока, что доказывается массовым заражением головных и платяных вшей и человеческих блох.
Мы считаем ошибочным мнение некоторых эпидемиологов начала XX в., что причиной всех или большинства эпизоотии среди сурков (Подбельский, 1901; Дудченко, 1909) и домовых мышей (Кольцов, 1915) служило заражение зверьков от трупов погибших от чумы людей. Однако отрицать возможность заражения грызунов при обгрызании ими человеческих трупов видимо не следует. Необходимо напомнить, что количество умерших при чумных эпидемиях в прошлые века было необычайно велико. Например, в пандемию XIV в. в Китае погибло до 13 млн., в других странах Востока - 24 млн., а в Европе вымерла четвертая часть ее населения - 25 млн. человек (Федоров, Рогозин, Фенюк, 1955). Учитывая, что захоронение трупов практически не осуществлялось, можно себе представить объем контакта с ними грызунов, как сожителей человека, так и обитателей естественных биоценозов.
Видимо значительно чаще алиментарного пути передача возбудителя чумы от человека к грызунам осуществлялась трансмиссивным путем. Блоха человека (Pulex irritans L) до недавнего прошлого во многих районах была обычнейшим паразитом в жилищах. В примитивных строениях с земляным полом численность блох достигала колоссальных размеров (Касуга и Танимото, 1941; Юркина, 1949). Эпидемиологическое значение этого вида достаточно велико (Блан и Балтазар, 1941, 1943; Новикова и Лала-заров, 1944). Эксперименты, поставленные А. А. Флегонтовой и Л. С. Малофеевой (1957), показали, что человеческая блоха легко заражается возбудителем чумы, длительно сохраняет его и хорошо передает укусом. Инстинными хозяевами P. irritans являются хищные мелкопитающие, но она чрезвычайно активна, подвижна и легко переходит на самых разнообразных животных. В северо-восточном Китае эту блоху регулярно обнаруживали на серой крысе ном Китае эту блоху регулярно обнаруживали на серой крысе (Абе, 1942; Касуга и Танимато, 1941). И. Г. Иофф (1949) указывает, что в ряде местностей на Тянь-Шане человеческая блоха в большом количестве встречается на сурках и в их норах.
По данным В. М. Попугайло (1959), в одном из районов Внутренней Монголии P. irritans составляла 18% от всех блох, паразитировавших на даурских сусликах. Р. Хорхе (1933), Г. Бланк и М. Балтазар (1943) указывают, что в эпидемии прошлых веков трансмиссивный обмен возбудителем от грызуна к человеку и обратно от человека к грызуну был нормальным явлением. Анализируя закономерности существования чумных очагов Китая, Б. К. Фенюк (1959, стр. 15) пишет: «В юго-восточных провинциях большое значение имела передача чумы человеческими и крысиными блохами от больного человека крысам, как один из факторов длительного сохранения «крысиной» чумы. Применение здесь после 1949 г. современной системы мер по локализации и ликвидации вспышек чумы прервало передачу возбудителя как внутри популяции крыс, так и от человека к крысам».
Все изложенное позволяет считать, что трансмиссивная передача возбудителя чумы от человека к грызунам относительно проста и вероятно в прошлом неоднократно осуществлялась. Еще проще возбудитель чумы попадал за пределы своего естественного ареала, когда человек со своими транспортными средствами невольно развозил по земному шару больных чумой синантроных грызунов. В литературе описано большое число случаев проникновения возбудителя чумы этим путем в чуждые для него ландшафтные условия (Федоров и Козакевич, 1957).
Несомненно, становление современного ареала чумного микроба, в первую очередь, определялось формированием ареалов его основных носителей и переносчиков. Детальному рассмотрению этого процесса мы посвятили специальную работу (Кучерук, 1965). Однако некоторые авторы считают, что на многие участки земной поверхности - «потенциальные очаги» возбудитель чумы не смог распространиться естественным путем. Наиболее четко эту точку зрения сформировал Ю. М. Ралль (1944, стр. 265): «В одних потенциальных очагах факторы очаговости также имеют место и хорошо выражены. В случае заноса сюда чумного вируса в достаточном количестве здесь возникают действующие очаги, отсутствие чумы в этих местностях определяется исторической случайностью, иногда - наличием временного барьера для проникновения инфекции».
Неясно, существовали ли к началу антропогена на территории Евразии «потенциальные» природные очаги чумы. Однако, несомненно, что на протяжении последних тысячелетий, человек, помимо своего желания разносил чумного микроба почти по всей территории этого материка. Возбудитель чумы мог неоднократно переходить от человека в популяции местных грызунов и закрепиться (акклиматизироваться) там, где ему позволяли сделать это существующие экологические условия. Следовательно, ареал возбудителя чумы (область распространения природных очагов ее) на Евразийском материке близок к потенциально возможному. Существование здесь «потенциальных природных очагов», как их понимает Ю. М. Ралль, кажется нам маловероятным.
В последние десятилетия появилось значительное количество работ, посвященных описанию территориальной приуроченности и эколого-географической характеристики отдельных природных очагов чумы и общему анализу географического распространения природных очагов - ареалу возбудителя чумы. Было установлено, что на Евразийском материке ареал чумного микроба (природные очаги чумы) лишь незначительно выходит к северу за пределы 50° с. ш., достигая 50°40' в Забайкалье и 51° на западном берегу р. Урала. На всем остальном пространстве северная граница природных очагов лежит южнее 50° с. ш. (Иофф, 1941). Мировое распространение природных очагов чумы и более детально очаги СССР и прилежащих стран были рассмотрены Ю. М. Раллем (1958).
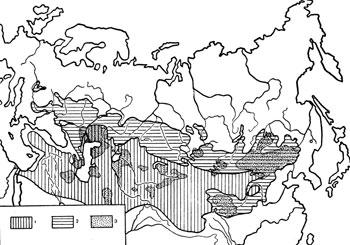
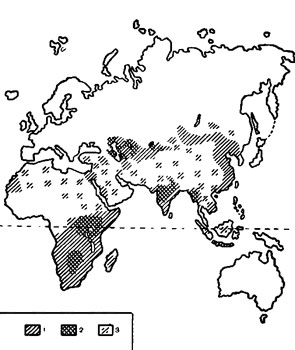
Мы попытались.провести анализ зональной приуроченности природных очагов чумы и разобрать факторы, которые могут обусловливать их распространение во внетропичесокй Евразии. С этой целью, на основании литературных источников (Геоботаническая карта СССР, 1956; Lesnicky a myslivecky atlas, 1955; Ковда, 1959), была составлена карта распространения степей и пустынь внетропической Евразии и на нее нанесены все достоверно выявленные в настоящее время природные очаги чумы (Мы вполне согласны с В. Н. Федоровым и В. П. Козакевичем (1957) в том, что громадные пустынные пространства Малой Азии, Иранского нагорья, Афганистана и Западного Китая недостаточно исследованы, и там, вероятно, будет еще выявлен ряд природных очагсв чумы, однако эти «невыявленные» очаги вряд ли существенно изменят очертания ареала возбудителя чумы во внетропической Евразии ) (рис. 1). При рассмотрении карты видно, что во внетропической Евразии природные очаги чумы не выходят за пределы степной и пустынной зоны. В то же время на карте распространения природных очагов чумы, опубликованной Ю. М. Раллем (1958), область распространения возбудителя чумы показана охватывающей почти всю Азию, лежащую к югу от границ СССР. На этой карте область распространения природных очагов чумы доходит до тихоокеанского побережья на отрезке от северной части Корейской Народно-Демократической Республики до провинции Гуандун (КНР) на юге (рис. 2), т. е. согласно приводимой Ю. М. Раллем карте, природные очаги чумы на востоке внетропической Евразии широко распространены в лесных областях: Дальневосточной хвойно-лесной и Японо-Китайской субтропических вечнозеленых лесов (Лавренко, 1950). На самом деле, восточные пределы распространения природных очагов чумы в умеренном и субтропическом поясе Евразии совпадают с областью доминирования фаунистического комплекса степных млекопитающих (Кучерук, 1959) и не переходят восточных границ степной зоны.
Интересно, что в западной части Евразийского материка природные очаги чумы почти не выходят за пределы пустынной зоны, лишь незначительно проникая в подзону пустынных степей (Волго-Донское междуречье и правобережье р. Урал). В восточной части материка, наоборот, основная площадь природных очагов чумы лежит в степной зоне. Очаги здесь доходят до северных и восточных границ степной зоны, широко проникая в пределы лесостепи (рис. 1). Таким образом, видно, что ареал возбудителя чумы в Евразии не обнаруживает строгой приуроченности к одной ландшафтной зоне, а связан с группой последних. Его северная граница на отдельных участках совпадает с северной границей степной зоны, а на юге он уходит в пределы тропических пустынь.
Необходимо рассмотреть, насколько ареал чумного микроба связан и зависит от распространения его теплокровных хозяев. По этому поводу существуют две противоположные точки зрения. Ю. М. Ралль (1957, стр. 115) пишет: «Общие географические границы зоны природной очаговости чумы определяются ареалами постоянных носителей этой болезни». Б. К. Фенюк (1958), наоборот, подчеркивает, что географические границы природной очаго вости чумы не совпадают с границами ареалов носителей (источников) и переносчиков чумного микроба, т. е. определенных видов грызунов и их блох. Ареалы и тех и других значительно шире и, как правило, далеко выходят за границы природных очагов.
На территории внетропической Евразии к основным носителям чумы относятся представители двух семейств грызунов: беличьих (Sciuridae) и хомякообразных (Cricetidae). Среди беличьих в роли основных носителей чумы выступают суслики и сурки. В семействе хомякообразных к основным носителям чумы относятся некоторые виды подсемейства песчанок, представители рода Merio-nes и Rhombomys.
Сопоставление ареала евразийских сурков (род Marmota) с областью распространения природных очагов чумы в Евразии (рис. 3) показывает их большое несоответствие. На значительном пространстве к западу от 70° в. д. ареал сурков лежит севернее области распространения природных очагов чумы. Восточнее этого меридиана ареал сурков совпадает с этой областью на большой площади. Но и здесь сурки распространены намного дальше к северу и, по-видимому, несколько южнее области распространения природных очагов, а восточные границы последней проходят значительно восточнее границы ареала сурков.
Из 6 видов сурков, обитающих в Евразии, ареалы 3 видов лежат за пределами природных очагов и у 3 видов лишь меньшая часть ареала совпадает с природными очагами чумы. У степного сурка (М. bobac Mull), имеющего 5 хорошо выраженных подвидов, ареалы двух из них лежат за пределами области распространения природных очагов чумы, у одного - большая часть ареала лежит за ее пределами и у двух подвидов основная площадь ареала занята природными очагами (табл. 1). Характерно, что ни у одного из сурков нет полного или почти полного совпадения ареала с территорией, занятой природными очагами.
Природные очаги чумы не распространились на весьма значительную Приуральскую и Северо-Казахстанскую часть ареала степного сурка. Поселения сурков в этих местах достаточно обширны, а плотность населения зверьков довольно велика. Трудно предполагать, что природные очаги чумы здесь не выявлены, гак как местные сурки издавна являются объектом промысла, интенсивность которого особенно повысилась с начала нынешнего века. Широкое земледельческое основание этих территорий началось лишь несколько лет тому назад и, следовательно, не могло сказаться на затухании очагов до самого последнего времени.
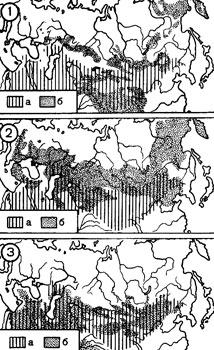
Большая часть ареала сусликов (род Citellus) в Евразии лежит севернее области распространения природных очагов чумы. На западе распространение этих зверьков также простирается на значительное расстояние за пределы природных очагов (рис. 3). Из 6 видов сусликов, обитающих в Евразии, ареалы двух видов целиком лежат за пределами области распространения природных очагов чумы и у трех видов они лишь незначительной частью заходят в эту область. Ареалы трех видов наполовину совпадают с областью распространения природных очагов и у одного вида совпадают почти целиком (табл. 1).
| Вид или подвид | Степень совпадения ареала с природными очагами ( |
Эпизоотическая значимость вида ( |
|---|---|---|
| Marmota marmota L. | --- | - |
| Marmota camtschatica Pall. | --- | - |
| Marmota menzbieri Kaschk. | --- | - |
| Marmota caudata Geoffr. | +- | ++ |
| Marmota himalayana Hodg. | +- | + |
| Marmota bobac Mull. | +-- | ++ |
| Marmota bobac Mull. | --- | - |
| Marmota schaganensis Bach. | --- | - |
| Marmota baibacina Kastsch. | +-- | - |
| Marmota centralis Thorn. | ++- | ++ |
| Marmota sibirica Radde. | ++- | ++ |
| Citellus sitellus Z. | --- | - |
| Citellus suslicus Guld. | --- | - |
| Citellusmajor Pall. | --+ | + |
| Citellus intermedius Br. | -+ | + |
| Citellus relictus Kaschk. | --+ | - |
| Citellus pygmaeus Pall. | -+ | ++ |
| Citellus dauricus Br. | -+ | ++ |
| Citellus fulvus Licht. | -++ | + |
| Citellus undulatus Pall. | --+ | + |
| Brarachi ones przewalskii Buch. | ? | - |
| Rhombomys opimus Licht. | +++ | ++ |
| Meriones meridianus Licht. | +++ | ++ |
| Meriones libicus Licht. | +++ | ++ |
| Meriones unguiculatus M. - Edw. | -++ | ++ |
| Meriones tamariscinus Pall. | +++ | + |
| Meriones persicus Blanf. | -++ | + |
| Meriones blacleri Thorn. | -++ | + |
| Meriones vinogradovi Hept. | +++ | - |
| Meriones crassus Sund. | -++ | - |
Интересно, что в некоторых случаях область распространения природных очагов не везде совладает с оптимумом ареала и участками наиболее высокой численности населения даже у видов, относящихся к основным носителям чумы. Малый суслик распространен от Днепра до 80° восточной долготы. На пространстве от левобережья Дона до низовьев р. Сары-су значительная часть ареала этого зверька совпадает с распространением природных очагов чумы. Севернее Аральского моря область распространения природных очагов чумы не достигает Мугоджар, хотя к последним приурочены наиболее древние поселения малого суслика, характеризующиеся весьма высокой численностью. По мнению С. Н. Варшавского (1962), именно к территории Мугоджар приурочен один из центров формирования современного ареала этого вида.
Во внетропической Евразии область распространения природных очагов чумы лишь незначительно выходит за пределы ареала подсемейства песчанок (Gerbillinae). Между Доном и Волгой и но среднему течению р. Урал природные очаги чумы простираются несколько севернее северо-западной границы ареала песчанок. В Забайкалье и Маньчжурии природные очаги занимают значительные площади.
У 10 видов подсемейства песчанок большая часть ареала или Еесь ареал лежат в пределах внетропической Евразии. Распространение всех этих видов на значительной площади совпадает с областью, занятой природными очагами чумы. Пять видов имеют ареалы, перекрытые природными очагами чумы наполовину и более а у других 5 видов ареалы полностью или резко преобладающей частью совпадают с областью распространения природных очагов чумы (табл. 1). Из 10 видов этого подсемейства, свойственных внетропической Евразии, 4 вида уже зарегистрированы в качестве основных носителей чумы в отдельных природных очагах.
Подытоживая вышеизложенное, следует отметить, что ареалы родов Citellus и Marmota в Евразии на значительном пространстве не совпадают с областью распространения природных очагов чумы (Такая же картина наблюдается в Северной Америке). Многие виды этих родов обитают севернее и западнее области распространения возбудителя чумы. Ареал подсемейства песчанок во внетропической Евразии, наоборот, несколько уже области распространения природных очагов чумы. На западе и особенно на востоке природные очаги чумы занимают значительные площади к северу от северной границы области распространения песчанок.
Следовательно,четкого совпадения ареала возбудителя чумы с ареалом родов и подсемейств млекопитающих, к которым относятся основные носители чумного микроба, в Евразии не наблюдается. Точно также не наблюдается совпадения ареала возбудителя чумы с ареалами основных переносчиков этой инфекции различных родов и видов блох (Иофф, 1941). Видимо ареал чумного микроба определяется какими-то специфическими биоценотическими отношениями, в которых немалую роль имеет прямое лимитирующее воздействие абиотических факторов.
Мы провели сравнение ареала чумного микроба во внетропической Евразии с распространением отдельных климатических показателей. Оказалось (рис. 4), что в пределах рассмотренной территории область распространения природных очагов чумы довольно близко совпала с границей годовой суммы осадков в 300-350 мм (на части материка, лежащей южнее 50° с. ш.). Некоторое проникновение чумных очагов за пределы этой границы наблюдается лишь в тех местах, где на небольшом протяжении резко меняется средне-годовое количество осадков (на крайнем востоке ареала чумного микроба).
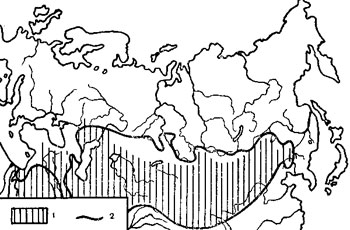
Еще более близкое совпадение границ ареала возбудителя чумы наблюдается с изолиниями определенного гидротермического коэффициента (Отношение суммы осадков к сумме температур, уменьшенное в 10 раз) - условного показателя, используемого агрокли-матологами (Селянинов, 1930; Сапожникова, 1956) для характеристики влагообеспеченности территории (рис. 5).
Связь природных очагов чумы с засушливыми территориями видимо объясняется следующим. В норах грызунов, лишенных хозяина, сухость позволяет гнездам сохраняться без гниения весьма продолжительное время (до нескольких лет). Этим создаются условия для длительного переживания эктопаразитов (преимущественно блох) и сохранения последними возбудителя чумы в отсутствии хозяина (Миронов, 1958). Вероятно, именно возможность сохранения возбудителя чумы после гибели владельца норы до времени нового заселения ее теплокровным хозяином и обеспечивает устойчивую циркуляцию инфекции на определенном участке земной поверхности.
Уже в 30 годах советскими учеными (Туманский и Поляк, 1931; Тинкер и Ступницкий, 1932; Тинкер, 1940) было установлено сохранение зараженных чумой блох малого суслика (Neopsylla setosa) в норах без зверьков на протяжении 5-8 месяцев. В замурованных, нежилых норах южно-африканских песчанок (Tatera brantsi), зараженные чумой блохи (Xenopsylla philoxera) сохранились в течение 120 дней (Девис - Davis, 1953), а в лишенных обитателей норах серого сурка, зараженные чумой Rhadinopsylla li были обнаружены через 415 и 417 дней (Шарец и др., 1958).
Если согласиться с гипотезой, высказанной в последнее время Г. Молларе, Ю. Карими, М. Ефтекари и М. Балтазаром (Н. Mol-laret, J. Karirni, M. Eftekhari et M. Baltazard, 1963) о длительном сохранении возбудителя чумы в почве глубоких нор, то лимитирующее значение определенного гидротермического режима почвы выступает еще яснее.
Видимо, настало время для проведения детальных, сравнительно-экологических исследований внешне сходных очаговых и неочаговых территорий. Именно сравнительным путем мы скорее всего сможем нащупать те тонкие механизмы, которые обусловливают саму возможность существования феномена природной очаговости на определенном участке земной поверхности.
Литература
Абе. Чума в Маньчжоу-го. «Bull, scientific res. Manchoukuo», 1942, 6, № 3, 2-19 (на японском языке).
Балабакин А. К., Солодкая А. Д. Восприимчивость к чуме когтистых песчанок при экспериментальном заражении. В. кн.: «Изв. Иркутского н.-и противочумного ин-та Сибири и Дальнего Востока», 1956, т. 14, стр. 19-25. 264
Балтазар М. Стойкость чумы в постоянных очагах. «Журн. гиг. эпидем., микробиол. и иммун.». 1964, 8, № 4, 333-343.
Беклемишев В. Н. Некоторые принципы нозогеографии облигатно-трансмиссивных болезней, поражающих человека. «Мед. паразитол. и паразитарные болезни», 1959, 28, № 6, 648-657.
Варшавский С. Н. Возрастные типы поселений и история расселения малого суслика. В кн.: «Исследования географии природных ресурсов животного и растительного мира». Изд. АН СССР М., 1962, 59-79.
Геоботаническая карта СССР, масштаб 1:4000000, под редакцией Е. М. Лавренко и В. Б. Сочавы, М.-Л., 1956.
Дербек Ф. А. История чумных эпидемий в России с основания государства до настоящего времени. СПБ, 1905.
Дудченко И. С. Об исследовании чумных заболеваний в Забайкальской области в 1908 г. в связи с тарбаганьей чумой. «Вестн. общей гигиены», 1909, июнь, июль.
Жданов В. М. Заразные болезни человека. М., 1953.
Иофф И. Г. Вопросы экологии блох в связи с их эпидемиологическим значением. Пятигорск, 1941.
Иофф И. Г. Aphaniptera Киргизии. В кн.: «Эктопаразиты», 1949, вып. 1.
Калабухов Н. И. Значение грызунов как фактора очаговости некоторых инфекций. «Зоол. журн.», 1949, 27, вып. 5, 389- 406.
Касуга и Танимото. Распространение грызунов и их блох в связи с чумой в Маньчжурии. В кн.: «Тр. Дженцзятуньской противочумной станции», 1941, 1, вып. 1. (на японском языке).
Ковда В. А. Очерки природы и почв Китая. М., 1959.
Кольцов Г. И. «Врач, газета», 1915, № 17.
Кучерук В. В. Блохи Маньчжурии и их эпидемиологическое значение по данным японской литературы. В кн.: «Вопр. краевой, общей и экспер. паразитологии», 1949, т. 6, 5-15.
Кучерук В. В. Степной фаунистический комплекс млекопитающих и его место в фауне Палеарктики. В кн.: «География населения наземных животных и методы его изучения». Изд. АН СССР, М.; 1959, 45-87.
Кучерук В. В. Вопросы палеогенезиса природных очагов чумы в связи с историей фауны грызунов. В кн.: «Фауна и экология грызунов». М., 1965, вып. 7, 5-86.
Лавренко Е. М. Основные черты ботанико-географического разделения СССР и сопредельных стран. В кн.: «Пробл. ботаники». Изд. АН СССР, М.-Л., 1950, вып. 1, 530-548.
Лобанов В. Н. Патологическая анатомия и патогенез чумы у человека. М., 1956.
Миронов Н. П. О существовании в прошлом природной очаговости чумы в степях южной Европы. ЖМЭИ, 1958, № 8, 18-23.
Новикова Е. И., Лалазаров Г. А. К вопросу о роли человеческих блох в эпидемиологии чумы. «Вестн. микробиол., эпидемиол. и паразитологии». Сб. работ, посвященных 25-летнему юбилею ин-та «Микроб». Саратов, 1944, 81-82.
Павловский Е. Н. О принципах изучения географии при-родно-очаговых и паразитарных болезней. В кн.: «Материалы ко Еторому съезду Географического об-ва СССР», М., 1954, 89-118.
Подбельский А. И. Наблюдения над тарбаганами в Монголии. «Русск. Арх. патологии», 1901, т. 12.
Попугайло В. М. К эпидемиологии чумы в Центрально-Маньчжурском очаге. В кн.: «Изв. Иркутского противочумного ин-та Сибири и Дальнего Востока», Улан-Удэ, 1959, 20, 67-75.
Ралль Ю. М. Очаговость чумы на грызунах в свете эколого-географических представлений. «Зоол. журн.», 1955, т. 23, вып. 5, 258-266.
Ралль Ю. М. Лекции по эпизоотологии чумы. Ставрополь, 1958.
Сапожникова С. А. Агроклиматическое районирование территории освоения целинных и залежных земель. В кн.: «Освоение целинных и залежных земель в 1954 г.». М., стр. 144-151.
Селянинов Г. Т. К методике сельскохозяйственной климатографии. «Тр. сельско-хоз. метеорологии», 1930, вып. 22.
Скородумов А. М. О восприимчивости Micromys minutus Pall к экспериментальной чуме. В кн.: «Изв. противочумного ин-та Сибири и ДВК». М.-Иркутск, 1936, 3, 32-33.
Тарасов П. П., Абрамова С. Г., Демидова Е. К. Высокогорная полевка и некоторые наблюдения над ее восприимчивостью к чуме. В кн.: «Изв. Иркутского противочумного ин-та Сибири и Дальнего Востока». 1949, т. 7, 90-106.
Тинкер И. С. Эпизоотология чумы на сусликах. Ростсв-на-Дону, 1940.
Тинкер И. С., Ступницкий П. Н. К вопросу о сохранении чумы в блохах суслика в межэпизоотическом периоде. «Жури, эпидемиол. и микробиологии», 1932, № 11 -12, 80-84.
Тихомирова М. М. Песчанка полуденная (Meriones meri-dianus) носительница чумного вируса в песчаных районах южных Волжско-Уральских степей. «Вестн. микробиол., эпидем. и паразитологии», 1934, 13, вып. 2, 89-102.
Туманский В. М., Поляк И. М. О сохраняемости чумного вируса в организме блох гнезда суслика в межэпизоотическое время. «Вестн. микроб., эпидемиол. и паразитологии», 1931, вып. 4. т. 10, 325-326.
Туманский В. М. Микробиология чумы. Медгиз, М., 1948.
Федоров В. Н., Козакевич В. П. Современное распространение чумы в зарубежных странах. В кн.: «Научн. конф. по природной очаговости и эпидемиологии особо опасных инфекционных заболеваний. Тезисы докладов». Саратов, 1957, 295-402. 266
Федоров В. Н., Рогозин И. И., Фенюк Б. К. Профилактика чумы. Медгиз, М., 1955.
Фенюк Б. К. Вопросы географии природных очагов чумы. «Зоол. журн.», 1958, 37, вып. 7, 1958, 961-970.
Фенюк Б. К. Заметки о природных очагах чумы и противочумной работе в Китайской Народной Республике (по путевым впечатлениям), ЖМЭИ, 1959, № 10, 8-15.
Флегонтова А. А., Малафеева Л. С. Роль человеческой блохи Pulex irritans L. в передаче чумы (экспериментальное исследование). В кн.: «Научн. конф. по паразитол., эпизоотол., эпидемиологии и другим вопросам природной очаговости чумы. Тезисы докладов». Ставрополь на Кавказе, 1957, 45-48.
Цен-Ко-Сюн. Экспериментальное изучение механизма передачи чумы среди когтистых песчанок. «Шуй-цонкан». 1958, № 3, 24.
Шарец А. С., Берендяев С. А., Красикова Л. В., Тристан Д. Ф. Эпизоотологическая эффективность разового истребления сурков. В кн.: «Тр. Средне-Азиатск. н.-и. противочумного ин-та», Алма-Ата, 1958, вып. 4, 145-147.
Щекунов З. И. К вопросу об отношении полевки Брандта к экспериментальной чуме. «Изв. Иркутского н.-и. противочумного ин-та Сибири и Дальнего Востока», 1957, т. 15, 95-102.
Юркина В. И. Материалы к экологии Pulex irritans. В кн.: «Тр. ин-та зоологии АН Украинской ССР», Киев, 1949, т. 2, 94-108.
Blanc G., Baltazard M. Recherches experimentales sur la peste. L'infection de la puce de L'homme, Pulex irritans. «Compt. Rend. Acad. Sci.», 1941, 213, N 22, 813-816.
Blanc G., Baltazard M. Quelques remarques a propos du memoire de Girard, «Bull. Soc. Path, ezot.», 1943, 36, N 7-8, 208- 217.
Davis D. H. S. Plague in South Africa; a study of the epizootic cycle in gerbils (Tatera brantsi) in the Northern Orange free state, «J. Hyg», 1953, 51, N 4, 427-449.
Jorge R. Summa epidemiologica de la peste. Epidemies ancies-nes et modernes. «Bull. off. Inter. Hyg. Publ.», 1933, v. 25, N 3.
Lesnicky a myslivecky atlas, Praha, 1955.
Mollaret H., Karimi J., Eftekhari M. et Baltazard M. La peste de fovissement. «Bull. Soc. Path, exot», 1963, 56, N 6, 1186-1193.
Pollitzer R. Plague «Wed. Health Org. Monograph series», Geneva, 1954, N 22.
Wu-Lien-teh A. Treatise of pneumonic plague. Geneva, 1926.
Москва, Институт эпидемиологии и микробиологии им. Н. Ф. Гамалеи АМН СССР
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://sohmet.ru/ 'Sohmet.ru: Библиотека по медицине'