О комплексных ландшафтно-эпизоотологических исследованиях природных очагов кожного лейшманиоза на юге Туркменской ССР (В. М. Сафьянова, Ю. А. Дубровский, В. М. Неронов, Е. М. Белова, В. Н. Вьюков, П. Д. Гунин)
Трудности в разработке и осуществлении мероприятий по охране населения от заболеваний зоонозным кожным лейшманиозом в значительной мере обусловлены слабой изученностью особенностей эпидемического и эпизоотического процесса на территориях с различными природными условиями.
В самой общей форме специалистам известно, что на территориях различного облика и географического положения опасность заражения кожным лейшманиозом неодинакова (Латышев и др., 1953). Однако, причины этого явления неизвестны. Следовательно, мы должны стремиться к разработке типологии природных очагов этого зооноза и таким образом обосновать тактику эпидемиологического прогноза и районированную систему профилактических мероприятий.
Обработка эпидемиологических данных позволила П. В. Кожевникову (1941), а в дальнейшем Н. Ф. Родякину (1955, 1957, 1962) и Е. С. Поповой (1963) выделить на территории Туркмении несколько крупных районов (долины Мургаба и Теджена, подгорная равнина Копет-Дага и т. д.), различающихся по уровню заболеваемости. Но очевидно, что в пределах крупных районов, на которые делится область природной очаговости лейшманиоза указанными выше авторами, могут быть выделены и более мелкие участки с разной степенью эпидемиологической опасности.
Полностью обследовать огромные территории Средней Азии, которые привлекаются в настоящее время для хозяйственного освоения, очень трудно. Поэтому нужно стремиться к тому, чтобы по ряду наиболее доступных признаков-индикаторов заранее определять степень такой опасности. Но для этого необходимо хорошо знать особенности циркуляции возбудителя в природных очагах и причинные зависимости, связывающие индикационные признаки с величиной риска заражения людей. Методы подобных исследований в медицинской географии разработаны еще недостаточно. При их разработке особое внимание следует уделить методам комплексных исследований, позволяющим получить сравнимые материалы для ландшафтно-эпизоотологической характеристики различных элементов структуры природных очагов лейшманиоза, для разграничения отдельных природных очагов и их типизации.
Комплексные исследования экспедиции ИЭМ им. Гамалеи по изучению кожного лейшманиоза в юго-восточной Туркмении были обеспечены тесным сотрудничеством зоологов, паразитологов, эпидемиолога и ландшафтоведов. Необходимо было установить конкретные формы зависимости пространственной структуры природных очагов лейшманиоза от структуры ландшафтов изучаемой территории. Если имеется такая зависимость, то нам важно было знать, комплексы какого ранга - урочища или ландшафты - определяют основные черты пространственного распределения носителей и переносчиков лейшманиоза (грызунов и москитов).
Исследования велись по следующим основным направлениям: 1) изучение эпидемиологической ситуации на обследуемой территории, 2) изучение ландшафтов, их морфологической структуры и отдельных компонентов, 3) изучение распространения эпизоотии кожного лейшманиоза среди теплокровных и распределения больных зверьков по территории, 4) исследование территориальных закономерностей в размещении видовых комплексов москитов, сравнительного обилия видов и их зараженности лептомонадами.
В ходе работы была выработана общая схема организации подобных исследований, уточнены некоторые методические приемы, сбора материала и его обработки. Широко использовался картографический метод, при котором все данные, полученные разными, специалистами, наносились на карты, анализировались и обобщались с учетом ландшафтных особенностей местности. Исследования экспедиции велись в разных масштабах: на стационаре в долине Мургаба и во время автомобильных маршрутов по Теджено-Мургабскому междуречью. В настоящем сообщении рассматриваются методы работы только в стационарных условиях.
Стационар экспедиции общей площадью около 50 кв. км расположен в окрестностях поселка Имам-баба, в среднем течении р Мургаб. Выбор этого участка не случаен, т. к. долина реки Мургаб давно известна, как один из наиболее неблагополучных. в отношении зоонозного кожного лейшманиоза районов Туркмении (Латышев и Позывай, 1937; Гусев, 1938 и др.). За последние годы данные регистрации этого заболевания по населенным пунктам в районе работ экспедиции отсутствовали. Поэтому нами было предпринято специальное обследование населения. С помощью подворьых обходов установлено, что в поселке Имам-баба около 50% местных жителей, а в соседнем с ним поселке совхоза «Большевик» до 77% местных жителей переболело кожным лейшманиозом. Ежегодно среди детей и приезжих наблюдается свежие случаи заболеваний. По отношению ко всем осмотренным неиммунным лицам они составили в 1962 году - 9,7%, в 1963 году - 10,2%, в 1964 году - 12,1%. Эти данные свидетельствуют о том,, что издавна известный очаг до сих пор сохраняет свою активность,
На территории стационара представлены три ландшафта: 1) ландшафт грядово-бугристых песков, 2) долинный ландшафт среднего течения р. Мургаб, 3) ландшафт полого-волнистых песков, которые хорошо различаются по своему генезису, по характеру форм рельефа и по почвенно-растительному покрову. Велики отличия этих ландшафтов и в морфологии. Каждый из них состоит из разного набора более мелких природно-территориальных комплексов. Так, в долинном ландшафте могут быть выделены следующие комплексы: 1) урочище первой надпойменной терассы с луговыми почвами под тугайными лесами, 2) сложное урочище второй надпойменной террасы с культурно-поливными и заброшенными землями, 3) урочища курганов, 4) сложное урочище третьей надпойменной террасы, 5) урочища шлейфов коренных берегов. и 6) урочища склонов коренных берегов. Схематичное распределение природно-территориальных комплексов показано на рис. 2. Подробно на методах выделения ландшафтов, изучении их морфологии и составлении ланфшафтных карт мы не останавливаемся, т. т сни детально освещены в специальных руководствах (Исаченко,. 1961; Анненская и др., 1962; Видина, 1962 и др.).
На территории стационара встречаются все млекопитающие, вообще зарегистрированные как участники эпизоотии кожного лейшманиоза. Численность больших песчанок и тонкопалых сусликов во всех пустынных ландшафтах находится на высоком уровне; численность мелких песчанок за годы наших работ была невысокой (табл. 1). Больших песчанок было особенно много в 1962-1963 гг., в 1964 г. их численность уменьшилась. Обилие мелких песчанок, наоборот, увеличилось в 1963 и 1964 гг.
| Природно-территориальные комплексы | Показатели численности | ||||||||||
| Набор больших песчанок на 1 га | Обитаемых нор больших песчанок, в % | Больших песчанок в одной норе | Тонкопалых сусликов на 1 га | Зверьков на 100 ловушкосуток | Пересечений следов на 1 км пыльной дороги | ||||||
| Краснохвостых песчанок | Полуденных песчанок | Домовых мышей | Ушастых ежей | Песчанок рода Meriones | Мелких мышевидных грызунов | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Полого-волнистые пески | >2 | 85 | 4,2 | 0,1 | 2,3 | 7,0 | 0,3 | - | - | - | |
| Грядово-бугристые пески | 2,3 | 85 | 4,7 | 0,7 | 0,5 | 5,7 | 0 | - | - | - | |
| Долина реки Мургаб | |||||||||||
| Склон песков правобережья | >2 | 88 | 4,8 | 0,4 | 1,2 | 8,2 | 0 | - | - | - | |
| Склон песков левобережья | 2,5 | 76 | 4,7 | 0,7 | 1,0 | 5,2 | 0 | - | - | - | |
| Вторая терраса левобережья | 1-2 | >50 | 3-4 | 0,1 | 1,0 | 10,0 | 0 | - | - | - | |
| Третья терраса правобережья | 2,3 | 74 | 3,4 | 0,2 | 3,5 | 4,0 | 0 | 2,5 | 12,9 | 12,9 | |
| Тугаи и заросли бурьянов рядом с ними | >0,1 | >50 | 3-4 | 0 | 0 | 0 | 6,5 | 4,2 | 7,0 | 3,5 | |
| Поля правобережья | 0 | 0 | 0 | 0 | 0 | 0 | 14,0 | 3,0 | 1,5 | 22,5 | |
| Поля левобережья | 0 | 0 | 0 | 0 | 0 | 0 | 15,5 | - | - | - | |
| Населенный пункт | 0 | 0 | 0 | 0 | 0 | 0 | 4,0 | 0 | 0 | 0,8 | |
Зараженность млекопитающих лейшманиозом устанавливали по наличию поражения кожи на участках головы с редким волосяным покровом (пораженными, как правило, бывают ушные раковины и очень редко веки, тубы или нос). Из пораженных участков делали мазки. При исследовании больших песчанок мазки, как правило, изготовляли из краевого участка ушных раковин всех зверьков - и больных, и внешне здоровых. При этом мы придерживались техники изготовления и фиксирования мазков, описанной Л. Н. Елисеевым и др. (1963).
Считается, что в эпизоотиях кожного лейшманиоза, кроме больших песчанок, участвуют краснохвостая и полуденная песчанки, тонкопалый суслик и ушастый еж (Латышев и др., 1953 и др.). Мы находили больных зверьков только среди больших песчанок (исследовано 2865 зверьков). Среди остальных грызунов (исследовано 97 краснохвостых песчанок, 198 полуденных песчанок, 120 сусликов) не было зверьков с внешне заметными поражениями кожи.
Поражения, сходные с лейшманиомами, обнаружены На ушных раковинах 4 ежей (всего исследовано 21), но в мазках из лих паразиты не обнаружены. Характерно, что среди этих так называемых второстепенных видов носителей мы не находили больных даже на участках весьма интенсивных эпизоотии, где зараженность больших песчанок превышала 20%.
Таким образом, показателем напряженности эпизоотической ситуации может служить количество больных особей в популяции больших песчанок. В результате анализа данных о территориальном распределении больных зверьков установлено, что места наиболее интенсивных эпизоотии приурочены к долинному ландшафту. Как видно на схематическом профиле (рис. 1), общее, количество больных песчанок выше.всего на террасах и по шлейфам коренных берегов, в песках правобережья оно минимально, а в песках левобережья интенсивность эпизоотии постепенно уменьшается с удалением от долины. При этом закономерно изменяется соотношение количества хорошо выраженных типичных леишманиом и мало заметных, нетипичных поражений ушной раковины. Как видно по рисунку (рис. 1 Б), количество больных песчанок с явными лейшманиомами выше всего на террасах и по кромке песков в долине; в песках правобережья оно минимально, а в песках левобережья значение показателя постепенно уменьшается с удалением от долины. Количество песчанок с сомнительными лейшманиомами почти везде находится на одном уровне и подобной закономерности не подтверждает. Таким образом, территориальные различия в степени интенсивности эпизоотии среди песчанок определяются, в основном, количеством больных особей с явными лейшманиомами. Тем самым для суждения о самых общих закономерностях распределения эпизоотии возможно использование результатов зи-зуального осмотра добытых зверьков. Это намного ускоряет работу, но, естественно, для проведения такого осмотра необходимы определенные навыки.
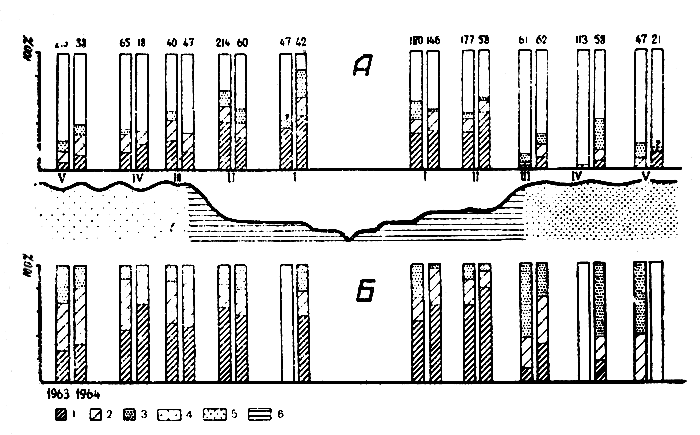
Рис. 1. Результаты изучения зараженности больших песчанок кожным лейшманиозом на территории стационара. А. Уровень зараженности; Б. Соотношение разных форм обнаружения лейшманий у зверьков. 1 - клинически выраженная форма; 2 - слабо клинически выраженная форма; 3 - без клинических проявлений; 4 - ландшафт грядово-бугристых песков; 5 - то же, полого-волнистых песков; 6 - то же, долины среднего течения р. Мургаб
Отмеченная ландшафтная приуроченность эпизоотических процессов разной интенсивности, по-видимому, является постоянной, повторяющейся ежегодно, т. к. и в 1963 году, и в 1964 году мы получили примерно одинаковые картины распределения больных песчанок по участкам стационара. Характерно, что эту общую закономерность не нарушают даже резкие изменения тюгодных условий (зима и весна 1964 г. были необычайно влажными, тогда как предыдущие годы были засушливыми). Не менее важно то, что некоторые изменения интенсивности эпизоотии в отдельных ландшафтах, которые были подмечены, произошли по-разному в каждом ландшафте. А именно, в полого-волнистых песках правобережья интенсивность эпизоотии после влажного 1964 г. заметно увеличилась, в грядово-бугристых песках левобережья несколько сократилась, а в долине на одних участках сократилась, на других - увеличилась.
Характерность эпизоотии определенной интенсивности для каждого ландшафта прослеживается не только во времени, но и в пространстве: в большинстве пунктов исследования песчанок, равномерно рассеянных по территории каждого ландшафта, мы регистрировали эпизоотию одного уровня, типичного для данного ландшафта, закономерно повторяющуюся на серии профильных разрезов (рис. 2, в каждом пункте добыто 20-25 песчанок). При этом изменчивость заболеваемости в отдельных сложных урочищах долинного ландшафта не существенно отличается от заболеваемости в других урочищах этого ландшафта. То же самое можно сказать и о заболеваемости в полого-волнистых песках правобережья Мургаба. В грядово-бугристых песках левобережья, более низких и более влажных, чем пески правобережья, по их окраине, граничащей с долиной, так же почти на всех профилях прослеживается постепенное уменьшение интенсивности эпизоотии с удалением от долины.
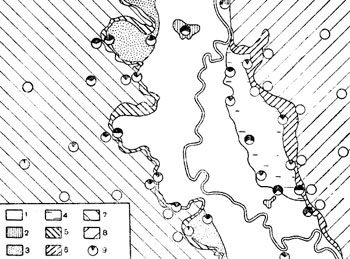
Некоторая изменчивость интенсивности эпизоотии в разных частях одного ландшафта, по-видимому, в ряде случаев имеет закономерный характер. Например, при крупномасштабном, исследовании распределения больных песчанок по отдельным колониям в пределах одного урочища выявлено, что зараженность зверьков в разных комплексах, составляющих это урочище, заметно отличается (рис. 3), поэтому можно считать, что различия в местной интенсивности эпизоотии в пределах сложного урочища обусловлены соотношением площадей участков с различной зараженностью песчанок.
Пользуясь теми же ландшафтными картами, мы провели наблюдения за москитами. Наиболее распространенный тип естественных убежищ и мест выплода москитов в изучаемых ландшафтах - это норы больших песчанок. Этот тип убежищ был выбран как основной объект для изучения пространственного размещения видовых комплексов москитов. Методика этого раздела работы состояла в следующем.
Ежегодно в каждом из основных урочищ долины и песков было под наблюдением 3-6 нор большой песчанки (всего 29 нор). В каждой норе в течение всего сезона активности москитов их еженедельно отлавливали жестяными воронками с садками из капрона на узком конце, установленными в 20 отверстиях каждой норы (экспозиция - 2,5 часа после захода солнца), а также на листы липкой бумаги, вставленные в 20 отверстий каждой норы (липучки держали в норах с 18 часов до 6 часов утра). В результате были получены данные об общих закономерностях распределения москитов.

В районе исследования встречаются 9 видов москитов: Phlebotomus papatasii. Ph. sergenti, Ph. caucasicus, Ph. andrejevi, Ph. mongolensis, Ph. chinensis, Sergentomyia arpaklensis, S. graecovi и S. clydei. На первом месте по численности стоит S. arpaklensis, субдоминантами являются Ph. papatasii и Ph. caucasicus. Остальные виды малочисленны. Общая численность москитов в долине заметно выше, чем в обоих песчано-пустыннных: ландшафтах. Особенно заметен резкий спад численности москитов при переходе от долинного ландшафта к ландшафту грядово-бугристых песков.
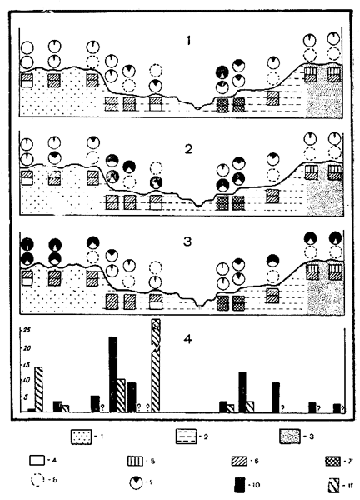
Рис. 4. Соотношение преобладающих видов Phlebotominae на разных участках стационара: 1 - Phlebotomus papatasii Sc, 2 - Sergentomyia arpaklensis Peri, 3 - Phlebotomus caucasicus Marz., 4 - Общая численность Phlebotominae. Условные обозначения: 1 - ландшафт грядово-бугристых песков; 2 - ландшафт долины Мургаба; 3 - ландшафт полого-волнистых песков; 4 - влажность грунта не исследовалась; 5 - влажность грунта до 2,5%; 6 - влажность грунта от 2,5 до 7,5%; 7 - влажность грунта более 7,5%; 8 - учеты москитов не проводились; 9 - процент данного вида от общего числа собранных Phlebotominae; 10 - общая численность москитов (среднее количество на 1 липучку) в 1964 г.; 11-то же, в 1963 г. Примечание: Процент москитов показан круговыми диаграммами: нижний ряд - 1963 г., верхний ряд - 1964 г., влажность грунтов показана прямоугольниками: верхний ряд - 1963 г., нижний ряд - 1964 г
Приуроченность различных видовых комплексов москитов к определенным ландшафтам и урочищам наиболее четко выявляется при анализе данных о пространственном распределении отдельных видов (рис. 41,2,3). Ph. papatasii - типичный обитатель долинного ландшафта. Как видно из рисунка, степень его доминирования выше всего в центральных урочищах долины. S. arpaklensis также специфический обитатель долины. Однако, он распространен в этом ландшафте более равномерно. Наоборот, Ph. caucasicus - доминирующий вид песчано-пустынных ландшафтов. В долине он встречается в урочищах третьей террасы и шлейфов песков коренного берега, где, однако, значительно уступает по своему обилию S. arpaklensis.
Изучение некоторых физико-химических свойств почво-грун-тов основных урочищ (см. ниже) позволило выявить несомненную связь степени доминирования отдельных видов москитов с влажностью грунта (рис. 4). Так, Ph. papatasii в своем распространении приурочен к урочищам с более влажным грунтом, причем наибольшее обилие этого вида выявлено на второй и третьей террасах правобережья, отличающихся самой большой влажностью (до 10%). Наоборот, для Ph. caucasicus выявлена обратная зависимость степени его обилия от влажности грунта. По-видимому, для этого вида более благоприятна влажность, свойственная песчаным ландшафтам.
Более или менее равномерное распределение S. arpaklensis во всех урочищах долинного ландшафта позволяет предположить, что все колебания влажности почво-грунтов долины лежат в пределах оптимума для этого вида. Однако на границе с песчаными ландшафтами, где существенно меняются показатели влажности, обилие S. arpaklensis заметно падает.
Резюмируя изложенное, можно сказать, что для комплекса видов москитов долины характерно доминирование Ph. papatasii и S. arpaklensis, для комплекса видов песчано-пустынных ландшафтов - доминирование Ph. caucasicus.
Эти общие закономерности пространственного распределения доминирующих видов москитов наблюдались как в засушливые 1962 и 1963 гг., так и во влажный 1964 г. (сопоставление данных 1963 и 1964. гг. см. на рис. 4). По-видимому, они остаются постоянными вне зависимости от метеорологических особенностей года. Нужно отметить при этом, что общая численность москитов в 1964 году по сравнению с 1963 годом в большинстве случаев возросла (рис. 44).
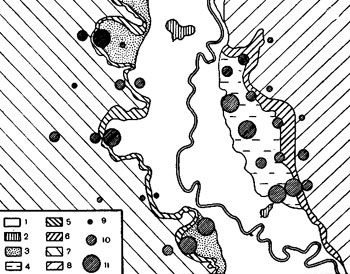
Особенности распределения видовых комплексов москитов и отдельных видов в пределах разных урочищ и ландшафтов мы изучили на той же серии профильных разрезов, которая была использована при изучении интенсивности эпизоотии среди больших песчанок. Москитов учитывали липучками одновременно в 120 колониях, по 4 в каждом пункте учета. Такие учеты проведены три раза в июле и в августе 1964 года. Эта работа показала неравномерность в распределении численности москитов в пределах каждого ландшафта. Однако, как видно из рис. 5, эта неравномерность не перекрывает основных особенностей распределения общей численности москитов по территории, которые установлены в результате вышеописанных наблюдений за контрольными колониями. Почти на всех профилях хорошо заметно снижение численности москитов при переходе от долины к пустынным ландшафтам.
Зараженность москитов лептомонадами мы изучали, во-первых, путем исследования мазков содержимого кишечника самок (более 1000 мазков) и, во-вторых, путем выделения живых культур лептомонад (около 600 посевов).
Судя по исследованию мазков, средняя зараженность Ph. рараtasii составила в 1963 г. 2,3%, в 1964 г. - 4,6%; S. arpaklensis, соответственно, 4,4 и 7,3%. Ph. caucasicus исследовались как в 1963, так и в 1964 г., но зараженные особи среди них выявлены только в 1964 году (2,6%). Таким образом, в 1964 г., по сравнению с 1963 годом, было отмечено некоторое увеличение зараженности лептомонадами всех доминирующих видов москитов.
Методом посевов на питательную среду (Белова и Сафьянова, 1963) исследовано 203 Ph. papatasii. От них выделено 8 культур лептомонад; от 295 исследованных S. arpaklensis выделено 20 кулыур лептомонад. Все посевы от Ph. caucasicus (80) были отрицательны. Зараженные москиты обнаружены на всех участках долины Мургаба, при этом нахМ не удалось выявить достоверной разницы в интенсивности зараженности лептомонадами москитов, населяющих отдельные урочища долинного ландшафта.
Значительная часть жизнедеятельности москитов проходит в норах большой песчанки, и, очевидно, здесь же происходит передача ими возбудителя лейшманиоза от одного зверька к другому. Для того, чтобы объяснить отмеченные выше ландшафтные отличия в зараженности больших песчанок и москитов и изменения этих показателей во времени, мы вынуждены были основное внимание обратить именно на норы больших песчанок. Оказалось, что строение нор и особенности их микроклимата в значительной мере задисят от их расположения и от особенностей почво-грунтов (плотности, механического состава, влажности грунта). Эти исследования позволили, в свою очередь, выявить определенную связь между изменением эпизоотической напряженности и изменением влажности почво-грунтов.
| Годы | Ландшафт грядово-бугристых песков | Долинный ландшафт среднего течения р. Мургаб | Ландшафт облесованных полого-волнистых песков | ||||||||
| Годы в 3 км от коренного берега | в 1 км от коренного берега | по кромке коренного берега | шлейф левого берега | 2-я терраса с песками | заброшенные земли на 2-й террасе | 3-я терраса | шлейф правого берега | по кромке коренного берега | в 1 км от коренного берега | в 3 км от коренного берегу | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1963 | 2,8 | 2,9 | 3,1 | 4,5 | ? | 13,0 | 12,8 | 3,3 | 1,7 | 1,9 | 2,2 |
| 1964 | ? | 2,7 | 2,9 | 2,5 | 2,9 | 10,6 | 11,9 | 5,1 | 4,5 | 5,5 | ? |
| Тенденция ( |
? | - | - | - | ? | - | - | + | + | + | ? |
Влажность почво-грунтов определялась нами с помощью ручного бура на глубине 1,5 метра по методу Н. В. Родионова и Н. И. Велрицкой (цит. по Добровольскому, 1962). Результаты двухлетних наблюдений за изменением влажности (более 200 измерений) в разных природно-территориальных комплексах представлены в табл. 2. Как отмечалось выше, 1963 г. был засушливым. Большое количество осадков зимой 1963-1964 гг. привело к заметному увеличению запасов влаги в облессованных песках правобережья и по шлейфу правого коренного берега. В то же время в остальных урочищах долинного ландшафта и в грядово-бугристых песках левобережья к осени, когда производились измерения, влажность почво-грунтов оказалась несколько ниже, чем в 1963 г. Наибольшее сокращение влажности отмечено на заброшенных землях второй террасы (на 2,4%) и на шлейфе левого коренного берега (на 2,0%). В обобщенном виде изменения влажности почво-грунтов в разных урочищах показаны на схематическом профиле (рис. 4). Если сопоставить с этим рисунком данные об изменении уровня зараженности больших песчанок (рис. 1А), то видно, что в полого-волнистых песках и на шлейфе правого коренного берега вместе с увеличением влажности почво-грунтов произошло увеличение интенсивности эпизоотии кожного лейшманиоза. На остальной территории стационара, за исключением второй террасы с навеянными песками и наиболее удаленных от долины участков грядово-бугристых песков, наблюдалось некоторое снижение интенсивности эпизоотии, и, как отмечалось выше, некоторое снижение влажности. Причины этих совпадений мы пока объяснить не можем. Однако они свидетельствуют о том, что метеорологические особенности зимне-весеннего сезона оказывают неодинаковое влияние на изменение интенсивности эпизоотии в условиях различных ландшафтов.
Результаты наших исследований показывают, что изученные в пределах стационара ландшафты различаются по всем признакам, определяющим напряженность эпизоотии: по зараженности больших песчанок, по характеру видовых комплексов москитов и их обилию, по зараженности москитов лептомонадами.
Следовательно, есть основание говорить о существовании на изученной нами территории трех самостоятельных природных очагов или трех хорошо выраженных морфологических частей природного очага зоонозного кожного лейшманиоза, приуроченных к каждому из изученных ландшафтов. Более точно определить их таксономическое положение мы пока не решаемся в связи с недостаточной разработанностью общих вопросов структуры природно-очаговых территорий. Окончательное определение ранга выделенных нами территорий требует дальнейших исследований.
Эти территории не равнозначны и по степени их эпидемиологической опасности: в долине реки постоянно регистрируется высокая заболеваемость людей, тогда как на Теджено-Мургабском междуречье, судя по нашим данным, случаи местных заражений отсутствуют.
Особенности структуры каждого природного очага (или его крупных морфологических частей) могут быть выявлены при изучении эпизоотических процессов, протекающих в урочищах и более мелких территориальных комплексах.
Возможно, что в других частях области природной очаговости кожного лейшманиоза будут выявлены какие-то иные закономерности существования природных очагов. Поэтому в заключение нам хотелось бы особенно подчеркнуть, что для того, чтобы все данные о разных типах природных очагов могли быть сопоставлены и сведены воедино, необходимо использование строго унифицированной методики подобных исследований. В этой связи мы надеемся, что изложенный нами опыт проведения стационарных работ может быть учтен при разработке такой методики.
Литература
Анненская Г. К., Видина А. А., Жучкова В. К., Коноваленко В. Г., Мамай И. И., Позднева М. И., Смирнова Е. Д., Солнцев Н. А., Цесельчук Ю. Н. Морфологическая структура географического ландшафта, Изд. МГУ, 1962.
Белова Е. М. и Сафьянова В. М. О методах изучения естественной зараженности москитов лептомонадами в очагах кожного лейшманиоза. «Зоол. журн.», 1963, т. 42, вып. 2.
Видина А. А. Методические указания по полевым крупномасштабным ландшафтным исследованиям. Изд. МГУ, 1962.
Гусев И. В. О заболеваемости кожным лейшманиозом в Мур-габском оазисе. «Сов. здравоохр. Узбекистана»,. № 4, 1938, 55-57.
Елисеев Л. Н., Сергиев В. П., Саакянц Э. М. К методике исследования больших песчанок на зараженность лейшманиозом. «Мед. паразитол. и паразитарные болезни», № 2, 1963, 223-224.
Исаченко А. Г. Физико-географическое картирование. Изд. ЛГУ, ч. 3, 1961.
Кожевников П. В. Кожный лейшманиоз в Туркмении и борьба с ним. В кн.: «Пробл. кожного лейшманиоза», Ашхабад, 1941, 93-100.
Добровольский В. В. Руководство к практическим занятиям по почвоведению. М., 1962.
Латышев Н. И. и Позывай Т. Т. Опыт эпидемиологического обследования в местах с кожным лейшманиозом в Туркмении. В кн.: «Проблемы паразитологии и фауны Туркмении». М.-Л.,. 1937, 163-181.
Попова Е. С. О лейшманиозах в Туркменской ССР и проблеме дальнейшего их изучения. «Здравоохр. Туркменистана», 1963, № 9, 29-33.
Латышев Н. И., Кожевников П. В., Повалишина Т. П. Болезнь Боровского. Медгиз, М., 1953.
Родякин Н. Ф. Болезнь Боровского (кожный лейшманиоз) в Туркменистане. В кн.: «Труды Туркменск. н-и кожно-венеролог. ин-та», Ашхабад, 1955, т. 4, 92-97.
Родякин Н. Ф. Болезнь Боровского в Туркменской ССР. В кн.: «Вопр. краевой патологии». Медгиз, М., 1957, 59-62.
Родякин Н. Ф. Кожный лейшманиоз (болезнь Боровского) в Туркмении и проблема его ликвидации. В кн.: «Вопр. краевой па-разитол. Туркмен. ССР». Ашхабад, 1962, т. 3, 63-70.
Москва, Институт эпидемиологии и микробиологии им. Н. Ф. Гамалеи АМН СССР; МГУ, Географический ф-т
Производственный календарь fastprint.info/regions/sankt-peterburg/calendar/.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://sohmet.ru/ 'Sohmet.ru: Библиотека по медицине'