Применение картографического метода при изучении размещения таежных клещей в природных очагах клещевого энцефалита (А. А. Земская, Л. Г. Суворова, Н. В. Туликова)
Значительное место в исследованиях очага клещевого энцефалита на юге Кировской области, проведенных эпидотрядом Института эпидемиологии и микробиологии им Н. Ф. Гамалеи АМН СССР, было отведено изучению роли таежных клещей (см.: Кучерук и др., настоящий сборник).
Изучение размещения клещей на территории очага складывалось из наблюдений за сезонным и суточным ходом активности и одномоментных учетов численности в большом числе точек. За изменением сезонной активности клещей следили на протяжении всех лет на двух, а в первые два года - на шести постоянных километровых маршрутах, расположенных в типичных биотопах. Учеты на них проводили каждые шесть дней по методике, предложенной во «Временных методических указаниях...» (1960). Суточную активность изучали на тех же маршрутах. Для этого периодические учеты на них повторяли каждые два часа.
Материал, характеризующий размещение клещей по территории, собирали во время сезонного пика активности (20 мая - 5 июня). Чтобы иметь сравнимые данные, учеты проводили почти одномоментно (за 5-7 дней) большой группой людей (около 20 человек). В часы максимальной суточной активности (17-20 часов) каждый сотрудник учитывал клещей на флаго-час в двух-трех заранее намеченных участках. Вследствие сильной захламленности и трудной проходимости территории клещей учитывали не на километр, а на флаго-час.
Планируя распределение мест учетов клещей, старались, чтобы вся территория стационара была достаточно полно покрыта учетами. Обычно на 1 квартал (2 кв. км) приходилось 6-10 учетов. И только в кварталах, где был обнаружен вирус, число учетных точек достигало 15. Количество учетов определялось характером растительности. Меньше учетов было в кварталах с однотипной растительностью, больше там, где характер растительности был более разнообразен. Места учетов выбирали по геоботаническим выделам. Каждое место учета лежало в пределах одного контура растительности. Пункты учетов взрослых клещей обычно совпадали с местами расстановки учетных линий для грызунов и повторялись из года в год. По возможности старались, чтобы эти учеты проводили одни и те же лица. Начинали учеты клещей с открытых вырубок, где пик активности клещей наступает раньше, затем учитывали в лесах.
Каждый учетчик получал картосхему участка, на котором он должен был провести учет. На эту же картосхему учетчик наносил полученные данные. Так, ежегодно мы получали сведения о численности клещей более чем со 100 участков нашего стационара и по ним составляли общую карту размещения взрослых клещей. Рабочей основой для ее составления служил крупномасштабный план местности.
Пользуясь вариационно-статистическим методом обработки данных, полученных при учете клещей, мы приняли четыре класса численности для взрослых клещей: I класс - от 1 до 3 клещей на флаго-час, II - от 4 до 9, III - от 10 до 15, IV - свыше 15. Схемы размещения наших объектов выполнены в двух вариантах - значковым методом и методом изолиний (рис. 1-5).
Анализ размещения взрослых клещей за все годы исследовав ния показал, что существуют годовые отличия в уровне численности и размещении клещей по территории. В годы обилия клещей пятна высокой численности охватывают большие площади, участки с нулевыми показателями почти не встречаются. В годы более низкой численности клещи размещаются мозаично, участки с нулевыми показателями подчеркивают пятнистость их распределения. Количество пятен высокой численности клещей и их положение на местности менялось по годам. Они постоянно перемещались по территории и только некоторые из них сохранялись на одном и том же месте, хотя границы их также менялись ежегодно.
Неравномерность распределения клещей на территории очага исследователи обычно связывают с характером биотопа, с про-кормителями предимагинальных фаз и взрослых клещей. Однако в оценке этих факторов у исследователей нет единого мнения. Одни утверждают, что образование участков повышенной численности таежного клеща определяется обилием крупных животных-про-кормителей взрослых клещей (Померанцев, 1935; Кузякин, 1942; Устинов, 1962 а, б). По мнению других, вокруг населенных пунктов создается зона более высокой численности таежного клеща за счет питания на домашнем скоте. В пределах же этой зоны неравномерность размещения клещей связана с неодинаковым обилием прокормителей личинок и нимф (Наумов и др., 1957; Шилова, 1959, 1961; Чабовский, 1959). Третьи считают, что ведущая роль принадлежит характеру биотопа, в частности, растительному покрову (Никифоров, 1959, 1960; Мишин. 1960; Мишин и Герасимова, 1962; Вершинский, Балаганов, 1962, а, б).
Картографический метод исследования позволяет довольно точно оценить значение каждого из вышеназванных факторов. Сопоставление карты растительности с картой размещения клещей показало, что в условиях нашего стационара очертания участков сходной численности не совпадают с границами биотопов. На рис. 1 показано, как пятна повышенной численности взрослых клещей в одних местах охватывают вырубки разного возраста и спелые сложные ельники, в других - пересекают однородные типы коренных лесов или вырубок. Изменение расположения и границ пятен повышенной численности таежного клеща в разные годы (см. рис. 2 и 3) также свидетельствуют о том, что характер растительности не определяет размещения клещей.
Эти материалы показывают несостоятельность широко распространенного мнения о том, что одинаковые лесотаксационные или геоботанические выделы имеют сходную численность клещей. Основанная на этом представлении методика учета клещей в ограниченном числе выделов и затем широкая экстраполяция средних данных на большую территорию при крупномасштабном и средне-масштабном картировании (Мишин, 1960), дает несоответствующую действительности картину размещения клещей.
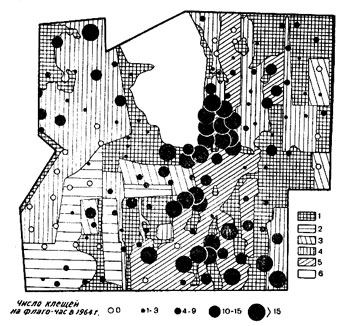
Размещение клещей имеет собственную структуру, не укладывающуюся в контуры биотопов, выделенных по признакам растительного покрова. К аналогичным выводам пришли Н. Б. Бируля и Л. И. Залуцкая (1961), Л. И. Залуцкая (1962), картографировавшие размещение клещей в очаге клещевого энцефалита на Урале.
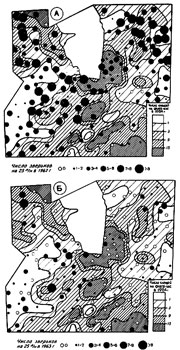
Для того чтобы оценить роль мелких млекопитающих в размещении клещей, проанализированы следующие материалы. Сопоставлено размещение численности взрослых клещей и мелких млекопитающих за годы, когда на зверьках прокармливались личинки и нимфы, давшие рассматриваемое поколение взрослых клещей. Такое сопоставление сделано для двух серий лет. На фоне численности взрослых клещей в 1963 г. значками показано обилие мелких млекопитающих в 1961 г. (рис. 2А), когда на них прокармливались личинки, и в 1962 г. (рис. 2Б), когда на зверьках кормились нимфы. Аналогичным методом сопоставлено размещение взрослых клещей в 1964 г. с обилием мелких млекопитающих в J962 г. (рис. ЗА) и в 1963 г. (рис. ЗБ). Все карты численности: грызунов относятся к началу июня (в 1961 г. - июню и июлю), т е. ко времени пика паразитирования предимагинальных фаз развития таежного клеща (Суворова и др., 1963).
В 1961 г. средне-взвешенная численность зверьков в июне составляла около 2, в июле - 7 зверьков на 100 ловушко-ночей (0,5 и 1,7 на учетную линию). По территории они распределялись неравномерно, места, где на учетные линии попадались зверьки, располагались отдельными пятнами среди площадей с нулевыми показателями улова (рис. 2А). В июне 1962 г. было 20 зверьков на 100 ловушко-ночей (в среднем 5 на учетную линию) (рис. 2Б и ЗА). Грызуны распределялись относительно равномерно, учетных линий с нулевыми показателями отлова было мало, на большей части территории попадалось от 5 до 17 зверьков на линии. Благодаря этому, личинки и нимфы практически всюду имели достаточное количество хозяев для прокормления. В 1963 г. было 5 зверьков на 100 ловушко-ночей (1,3 на учетную линию). По площади примерно половина стационара имела нулевые показатели стлоЕа, на остальной части - было от 1 до 5 зверьков на линии и лишь в единичных местах их число доходило до 9. Численность абсолютно доминирующей среди насекомоядных обыкновенной бурозубки в первой декаде июля в эти годы составляла в 1961 г. - 1, в 1962 г. - 16, в 1963 г. - 1 зверек на 10 канавко-суток. Материалов для составления карт размещения насекомоядных мы не имеем (10 ловчих канавок, расположенных в разных биотопах и непрерывно работающих в течение каждого сезона, не могут дать сведений о территориальном размещении животных на 40 кв. км).
Участки высокой численности взрослых клещей в 1963 г. не совпадают с местами обилия зверьков ни в 1961 г., ни в 1962 г. (рис. 2). Такое несовпадение наблюдается и во второй серии лет 1962-63-64 гг. (рис. 3). По-видимому, при относительно высокой весенней численности грызунов и землероек, их размещению не принадлежит ведущая роль в формировании участков различного обилия клещей.
Крупные дикие и домашние животные-прокормители половозрелых клещей, по мнению ряда авторов (Наумов и др., 1957; Шилова, 1959, 1961 и др.), определяют лишь общую территорию очага, но не влияют на характер размещения клещей на самой территории очага.
Наличие карт размещения взрослых клещей за пять лет и данных за несколько лет по вьшасу и прогону скота близлежащих деревень, позволило подойти к оценке роли скота в неравномерном распространении клещей. Оказалось, что на территории стационара большинство участков высокой численности находится в местах выпаса или прогона скота. Так, центральный участок существовал с 1961 по 1964 гг. (рис. 1), здесь на протяжении четырех лет до 1960 г. постоянно выпасался скот (до 80 голов) близлежащей деревни. В юго-восточном углу стационара постоянно прогоняли скот (около 150 голов), а с 1961 г. его стали здесь пасти. Отдельные небольшие участки высокой численности клещей были здесь все годы наших наблюдений, а в 1964 г. они слились и стали занимать значительную часть территории (рис. 1). На юго-западе стационара скот (120-180 голов) выпасался на протяжении многих лет, и в 1961 г. мы обнаружили здесь пятно высокой численности клещей. В 1962 г. по снегу ранней весной этот район был обработан ДДТ, в результате чего численность клещей здесь резко упала и не превышала 5 клещей на флаго-час. В течение последующих лет, вплоть до 1964 г., численность клещей здесь так и не восстановилась.

Наиболее низкая численность клещей на территории стационара все три года была в его северо-восточной части, где число клещей в период максимума не превышало 5 на флаго-час, за исключением небольшого пятна, где численность клещей была несколько выше - до 15 на флаго-час. В этом районе скот не выпасается, а существование пятна повышенной численности связано, очевидно, с тем, что в этих местах (по данным учетов зоологической группы) наиболее постоянно держались зайцы.
Таким образом, полученные материалы и их анализ показали, что в формировании участков повышенной численности в данном очаге ведущая роль принадлежит прокормителям половозрелых клещей.
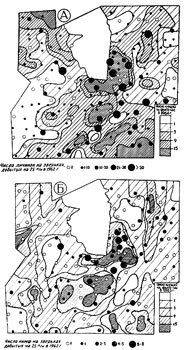
Мы попытались проследить формирование участков повышенной численности взрослых клещей на протяжении жизненного цикла. Для этого мы сравнили данные по территориальному распределен нию всех фаз развития в последовательности: нимфы -1962 г., взрослые клещи - 1963 г.; личинки - 1962 г., взрослые клещи - 1964 г. (В годы депрессии численности грызунов - в июне 1961 и 1963 гг. на большинстве учетных линий не были пойманы зверьки (рис. 2А и ЗБ) и, следовательно, невозможно было судить о наличии и численности личинок и нимф. Поэтому и нет карт размещения личинок 1961 и нимф 1963 гг) Анализируя карты размещения личинок 1962 г. и взрослых клещей 1964 г. (рис. 4А), нимф 1962 г. и взрослых клещей 1963 г. (рис. 4Б), можно отметить, что участки повышенной численности личинок обычно остаются участками повышенной численности взрослых клещей через год, так же, как участки обилия нимф почти все остаются участками повышенной численности взрослых клещей на следующий год. Следовательно клещи удерживались на данном участке в течение цикла развития вплоть до взрослой голодной фазы. Особенно отчетливо эта закономерность прослежена при изучении пораженности клещами птиц (Коренберг, и др., настоящий сборник).
Многолетние учеты личинок и нимф на зверьках, добытых постоянными ловчими канавками, показали, что индексы обилия личинок одного года всегда выше индексов обилия нимф следующего года. Но смертность нимф не одинакова в различных биотопах. На вырубках количество нимф, по сравнению с предшествующим количеством личинок, уменьшилось в 9-13 раз, в лесах - только в 2-4 раза. Очевидно, в липово-еловых лесах условия выживания предимагинальных фаз развития клеща значительно лучше, чем на вырубках разного возраста.
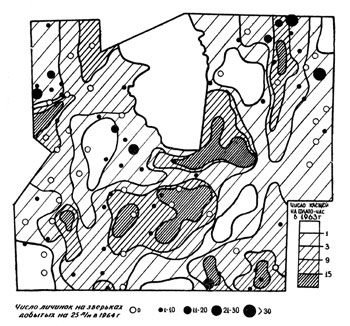
При сопоставлении размещения взрослых клещей предыдущего года с размещением личинок следующего года (рис. 5) наблюдается изменение в территориальном распределении клещей. Места обилия личинок чаще не совпадают с местами повышенной численности голодных взрослых клещей в прошлом году. Это также свидетельствует о том, что решающая роль в размещении клещей принадлежит питающимся самкам и соответственно их прокормителям (домашнему скоту, диким копытным, хищным и зайцам).
Роль прокормителей тгредимагинальных фаз совершенно иная. В очагах с относительно устойчивой высокой численностью грызунов и насекомоядных, зверьки поддерживают сложившееся размещение клещей до завершения цикла их развития. Там, где численность мелких млекопитающих испытывает длительные и глубокие депрессии, мозаичное распределение и отсутствие зверьков могут в одних местах ликвидировать, в других - сохранить запасы вылупившихся личинок и нимф. Но существенного перераспределения имеющихся клещей мелкие млекопитающие и птицы произвести не могут.
Аналогичную роль играют и условия метаморфоза всех фаз развития таежных клещей. В подходящих типах растительности личинки вылупляются из большинства кладок и перелинивает большинство напитавшихся личинок и нимф. Тем самым в благоприятных биотопах полностью сохраняется размещение напитавшихся самок и яйцекладок. В неподходящих типах растительности значительная или большая часть кладок и напитавшихся особей отмирает. Сочетание биотопов, в разной степени благоприятных для прохождения цикла развития таежных клещей, может лишь корректировать исходную численность клещей в разных местах.
Клещи удерживаются на данном месте в течение цикла развития от личинок до голодной взрослой фазы, а от напитавшейся отпавшей взрослой самки происходит изменение в их территориальном размещении. Это связано с выносом взрослых клещей крупными животными-прокормителями, которые разносят питающихся самок на значительные расстояния. Эти перемещения особенно сильно сказываются на дальнейшем распределении популяции клещей благодаря огромной плодовитости самок. Каждая перенесенная и выжившая самка дает начало тысячам особей клещей, тогда как перенос личинки или нимфы дает, в лучшем случае, только одну особь следующей фазы.
Исходя из вышеизложенного, становится понятным, почему территориальное распределение клещей в изучаемом очаге не совпадает с геоботаническими выделами и сохраняется резко неравномерным даже в годы, которым предшествует исключительное обилие грызунов, обеспечивающих прокармливание всего запаса личинок и нимф на всей территории. Таким образом, крупномасштабное картографирование фактической численности всех фаз развития клеща и основных прокормителей позволяет наиболее объективно оценить значение разных факторов в распределении таежных клещей на территории природного очага.
Литература
Бируля Н. Б., Залуцкая Л. И. К вопросу о внутриланд-шафтных очагах весенне-летнего клещевого энцефалита. В кн.: «Зоол. исслед. в природн. очагах клещевого энцефалита», М., 1961.
Временные методические указания по эпидемиологической, вирусологической, паразитологической работе в природных очагах клещевого энцефалита. Минздрав РСФСР. М., 1960.
Вершинский Б. В., Балаганов В. Я. Очаги таежного энцефалита и динамика растительного покрова. В кн.: «Докл. Ин-та географии Сибири и Дальнего Востока». Иркутск, 1962а.
Вершинский Б. В. и Балаганов В. Я. Опыт составления крупномасштабной медико-географической карты на ландшафтно-географической основе. В кн.: «Комплексное картографирование природы и хозяйства». Иркутск, 1962 б.
Залуцкая Л. И. К изучению Верхотурского очага клещевого энцефалита. «Вопросы экологии», Киев, 1962, 8.
Коренберг Э. И., Ковалевский Ю. В., Суворова Л. Г. Вопросы методики крупномасштабного картографирования при изучении роли птиц в природном очаге клещевого энцефалита. Наст, сборник.
Кучерук В. В., Земская А. А., Ковалевский Ю. В., Коренберг Э. И., Пчелкина А. А., Суворова Л. Г., Тупикова Н. В. Методика крупномасштабного картографического изучения пространственной структуры природных очагов болезней человека. Наст, сборник.
Кузякин А. П. О роли млекопитающих Уссурийского края в эпидемиологии клещевого энцефалита. «Зоол. журн.», 1942, 21, вып. 3.
Мишин А. В. Методика выявления потенциальных природных очагов клещевого энцефалита. Ижевск, 1960.
Мишин А. В. и Герасимова Е. Н. К вопросу картирования природных очагов клещевого энцефалита. В кн.: «Проблемы медицинской географии». Л., 1962.
Наумов Н. П., Шилова С. А., Чабовский В. И. Роль диких позвоночных в природных очагах клещевого энцефалита. «Зоол. журн.», 1957, 36, № 3.
Наумов Н. П. Теплокровные животные (преимущественно млекопитающие) как источник болезней человека. Тез. докл. Межинститутск. научн. конф. по заболев, с природн. очаговостью. Томский научн. иссл. ин-т вакцин и сывороток, Томск, 1956, 14.
Никифоров Л. П. Опыт зоолого-паразитологических исследований природных очагов клещевого энцефалита в Тяжинском районе Кемеровской области. Сообщ. 1. Природный очаг, его ландшафтная приуроченность и применение биоценологической методики исследования. «Мед. паразитол. и паразитарные болезни», 1959, № 6.
Его же. Опыт зоолого-паразитологического исследования природных очагов клещевого энцефалита в Тяжинском районе Кемеровской области. Сообщ. 2. Симморфология природных очагов (выявление биоценозов и их морфологическая характеристика) «Мед. паразитол. и паразитарные болезни». 1960, № 3.
Померанцев Б. И. К вопросу о происхождении клещевых очагов в Ленинградской области. В кн.: «Вредители животноводства». Изд. АН СССР, Л., 1935.
Суворова Л. Г., Туликова Н. В., Фарафонова Г. В. Роль различных мелких млекопитающих в прокормлении личинок и нимф таежного клеща в очаге клещевого энцефалита восточноевропейских южнотаежных лесов. В кн.: «Клещ, энцефалит и вирус, геомор. лихорадки. Материалы научн. конф.», Омск, 1963.
Чабовский В. И. Значение активных и пассивных перемещений клещей I. persulcatus в очагах клещевого энцефалита. В кн.: «IV съезд Всес. энтомол. о-ва. Тезисы докладов». М.-Л., 1959, 1.
Шилова С. А. Значение позвоночных животных в формировании очагов клещевого энцефалита. В кн.: «Десятое совещ. по паразитолог, пробл. и природноочаговым болезням». Изд. АН СССР. М.-Л., 1959.
Шилова С. А. Влияние экологических особенностей позвоночных животных на формирование очагов клещевого энцефалита. В кн.: «Зоол. исслед. в природн. очагах клещевого энцефалита». М., 1961.
Устинов С. К. К познанию экологии крупных млекопитающих в первичных очагах клещевого энцефалита. В кн.: «Тр. Иркутск, н.-иссл. ин-та эпидемиол. и микробиологии». Иркутск, 1962 а, 7.
Устинов С. К. Состояние и пути дальнейшего изучения роли крупных млекопитающих в очагах клещевого энцефалита на территории СССР. В кн.: «Тр. Иркутск, н.-иссл. ин-та эпидемиол. и микробиологии». Иркутск, 1962 б, 7.
Москва, Институт эпидемиологии и микробиологии им. Н. Ф. Гамалеи АМН СССР; МГУ, Географический ф-т
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://sohmet.ru/ 'Sohmet.ru: Библиотека по медицине'