Гормональная регуляция половой сферы ребенка на основных этапах развития
Взаимосвязь звеньев системы гипоталамус-гипофиз-гонады осуществляется на основе двойного саморегулирующегося процесса - отрицательной и положительной обратной связи. Характер этой взаимосвязи зависит от возраста и пола ребенка, поэтому комплексное состояние гипоталамических центров* и регулируемый ими уровень гипофизарных и половых гормонов принято обозначать термином "гонадостат" [Grumbach М.,. 19741.
Гонадостат функционирует на протяжении всего развития ребенка, начиная с внутриутробного периода. В процессе становления половой функции у ребенка можно выделить 4 основных "критических" периода: 1) внутриутробный (фетальный) 2) период новорожденности; 3) препубертатный; 4) пубертатный.
Фетальный период. В этом периоде гипоталамо-гипофизарно-гонадные взаимоотношения зависят от пола ребенка.
У мальчиков максимальное количество клеток Лейдига, продуцирующих тестостерон, зафиксировано между 10-й и 18-й неделями внутриутробного развития. В эти же сроки отмечена № максимальная концентрация тестостерона в крови плода, достигающая величин, свойственных взрослым мужчинам [Murrph I., 1974; Reyes F., 1974]. Физиологическое значение высокого подъема уровня тестостерона в этот период заключается прежде всего в участии тестостерона в формировании внутренних и наружных гениталий мальчика. В период формирования урогенитального тракта по мужскому типу и дифференцировки вольфовых протоков необходима высокая концентрация тестостерона и его активного метаболита 5α-дигидротестостерона. В противном случае урогенитальный тракт плода автоматически формируется по женскому типу.
Кроме того, активность фетальных яичек в указанный период обеспечивает половую дифференцировку гипоталамуса. Как известно, гипоталамическая регуляция гонадотропной функции имеет циклический характер у особей женского пола и постоянный, тонический характер у особей мужского пола. В работах ряда авторов показано, что половая дифференцировка гипоталамуса осуществляется в определенный период внутриутробной жизни и зависит от избытка половых стероидов [Бабичев В. Н., 1981]. Возможно, что 10-18-я недели внутриутробного развития плода с высоким подъемом концентрации тестостерона и есть один из "критических" периодов, обусловливающих половую дифференцировку гипоталамуса. Блокада циклического центра гипоталамуса состоит в утрате им способности вызывать подъем уровня ЛГ в ответ на воздействие пороговой концентрации эстрадиола (положительная обратная связь).
Тестостерон блокирует гипоталамус путем своего превращения в эстрогены. В гипоталамусе найдены рецепторы, активно связывающие тестостерон и андростендион, и ферменты, превращающие их в эстрадиол и эстрон.
Начальный этап стероидогенеза в фетальных яичках стимулируется материнским хорионическим гонадотропином, максимальная выработка которого наблюдается между 8-й и 12-й неделями беременности [Савченко О. Н., Степанова Н. А., 1979]. Затем по мере созревания морфологических структур гипоталамо-гипофизарной системы продукция андрогенов начинает регулироваться собственными гонадотропинами плода. Уровень ЛГ и ФСГ гипофиза плода мужского пола начинает нарастать с 12 нед внутриутробного развития, достигая максимума к 20-25-й неделе, затем постепенно происходит снижение уровня гонадотропных гормонов.
У девочек эндокринная активность фетальных гонад не является необходимой для половой дифференцировки внутренних и наружных гениталий. Дифференцировка ткани яичника начинается позже, чем ткани яичка. Первые ооциты появляются не ранее 12-й недели внутриутробного развития. Формирование примордиальных фолликулов достигает максимума между 12-й и 25-й неделей, к 26-й неделе можно отметить единичные зрелые фолликулы на стадии граафова пузырька [Forest M., Peretti E., 1976].
Продукция половых гормонов яичниками в период внутриутробного развития минимальна, и выделяется главным образом эстриол.
Яичники плода не способны продуцировать эстрогены в ответ на стимуляцию хориогонином. Стероидогенез в фетальных яичниках, видимо, стимулируется собственными гонадотропинами, которые гипофиз плода женского пола начинает продуцировать довольно рано-с 10 нед внутриутробного развития. Уровень ЛГ достигает максимума к 24-й неделе, а ФСГ - к 29-й неделе внутриутробного развития [Grumbach M., Kaplan S., 1973].
Содержание гонадотропных гормонов у плода женского пола значительно выше, чем у плода мужского пола. Возможно, это различие секреции гонадотропинов у плодов разного пола объясняется более ранним формированием обратной связи у мальчиков.
Период новорожденности. В периоде новорожденности происходит дальнейшая перестройка гипоталамо-гипофизарно-гонадных взаимоотношений. В течение первых дней после рождения из организма ребенка выводятся гормоны плацентарного происхождения, главным образом хорионический гонадотропин. Параллельно с этим происходит и резкое снижение уровня половых гормонов.
У мальчиков снижается уровень тестостерона. Однако уже через неделю после рождения он начинает постепенно повышаться и достигает максимума к концу первого месяца жизни. Высокая концентрация тестостерона держится 3 мес, достигает такого же значения, как в пубертате, а затем начинает снижаться [Forest M. et al., 1973]. В это время имеется большое содержание свободной фракции тестостерона. Физиологическое значение подъема содержания тестостерона в этот период не выяснено, но предполагают, что тестостерон продолжает "программировать" половую дифференцировку гипоталамуса, начавшуюся во внутриутробном периоде [De Moor P., 1973; Sizonenko P., 1975; Gustafson J., 1985].
У девочек подъем содержания эстрогенов, главным образом эстрадиола, приходится на конец второй недели жизни. Уровень эстрадиола у девочек значительно ниже, чем тестостерона у мальчиков, его выброс колеблется. Вместе с тем относительно высокий уровень эстрадиола у девочек держится дольше, чем тестостерона у мальчиков, - приблизительно в течение всего первого года жизни [Bidlingmajer R. et al., 1974]. Секреция гонадотропных гормонов гипофиза на первом году жизни довольно высокая у детей обоего пола, но уровень ФСГ у девочек значительно превышает этот показатель у мальчиков [Forest M., 1974].
Увеличенное количество гонадотропных гормонов у детей первого года жизни противоречит имеющемуся в этот период повышению уровня половых гормонов. Это противоречие объясняется незрелостью гипоталамических центров и гипофиза, контролирующих функцию гонад. Кроме того, в этом возрасте при незрелости гипоталамо-гипофизарных структур возможно существование биологически активных субъединиц ЛГ [Hagen С., McNeilli А., 1975].
Формирование взаимосвязей в цепи гипоталамус-гипофиз-гонады заканчивается к 11/2-2 годам. У ребенка 2-7 лет уровень гонадотропинов и половых стероидов низкий, но чрезвычайно высока чувствительность гонадостата. Кроме того, многие авторы предполагают воздействие в этот период центрального ингибирующего механизма, не зависящего от отрицательной обратной связи половых стероидов, который тормозит секрецию гонадотропинов и задерживает наступление пубертата [Escoibar М., 1976].
Препубертатный период (период адренархе). Качественные изменения гормональной сферы наступают после 6 лет, когда происходит созревание адреналовой системы с быстрым подъемом уровня надпочечниковых андрогенов: дегидроэпиандростерона и его сульфата и Δ4-андростендиона [Sizonenko Р., 1976; Reiter М., 1977; Apter D. et al., 1979].
Эта препубертатная фаза развития ребенка носит название адренархе. После наступления непосредственно пубертата уровень адреналовых андрогенов продолжает повышаться. Второй пик их подъема отмечается в 12-13 лет [Korth-Shultz S. et al., 1976]. Роль адренархе в организме до сих пор окончательно не выяснена.
Известны случаи быстрого полового развития при адекватном, но поздно начатом лечении детей с врожденной дисфункцией коры надпочечников. В связи с этим ряд авторов считают, что андрогены надпочечников, воздействуя на уровне гипоталамуса прямо или путем превращения в эстрогены, стимулируют созревание гипоталамо-гипофизарно-гонадной системы [Colli R., Duchatme J., 1975]. В то же время нормальное половое развитие лиц с первичной надпочечниковой недостаточностью или с ускоренным адренархе позволяет некоторым авторам считать, что надпочечниковые андрогены не влияют на сроки начала пубертата [Sizonenko Р., 1978].
Кроме роли надпочечниковых андрогенов в препубертате, остается неизвестным и фактор, индуцирующий их скачкообразный подъем. Никакого увеличения ЛГ, ФСГ и АКТГ при наступлении адренархе не отмечено [Apter D., 1979; Culter G., 1980]. Установлено, что уровень дегидроэпиандростерона не меняется при экзогенном введении АКТГ и хориогонина [Cutler J., 1980].
В последние годы появились сообщения о выделении гипофизарной фракции, специфически стимулирующей андрогенную зону надпочечников [Parker L. et al., 1979]. Возможно, эта oфракция обладает ЛГ-подобной активностью. В настоящее время в литературе обсуждается возможная роль пролактина как стимулятора адренархе. Это мнение подтверждается тем, что надпочечники имеют специфические рецепторы для пролактина, пролактин способен стимулировать продукцию Δ4-андростендиона in vivo, увеличивая активность 3β-ол-дегидрогеназы. Клинические исследования доказали большой подъем пролактина в ночные часы в препубертатный период [Parker L., Sach J., 1978; Sultan С., 1979].
Нам удалось подтвердить достоверное повышение уровня пролактина у здоровых девочек 6-8 лет (323±36,3 мед/мл), по сравнению с девочками пубертатного возраста (165±22,2 мед/мл; Р<0,05). У мальчиков подобной закономерности выявить не удалось. Вопрос о роли пролактина в процессе адренархе требует дальнейшего изучения.
Пубертатный период. Важнейшим этапом полового развития ребенка является пубертат. В этот период происходит сложная многоэтапная перестройка гипоталамо-гипофизарных взаимоотношений, она завершается репродуктивной зрелостью организма.
Основной гормональной характеристикой пубертата становится качественное изменение гипоталамо-гипофизарно-гонадных взаимоотношений в системе отрицательных и положительных обратных связей. По мере развития пубертата чувствительность гипоталамуса к половым стероидам постепенно падает и требуется все возрастающее количество половых гормонов для подавления выброса и секреции гонадотропных гормонов гипофиза. Снижение порога чувствительности гипоталамуса приводит к растормаживанию секреции гонадотропных гормонов, уровень которых постепенно нарастает.
Нарастание секреции гонадотропных гормонов гипофиза в течение пубертата неоднородно и зависит от пола ребенка. В начале пубертата отмечено значительное преобладание уровня ФСГ, особенно у девочек. Физиологическое значение этого факта, видимо, заключается в том, что ФСГ способствует повышению чувствительности ткани гонад к ЛГ, увеличивая тем самым секрецию половых стероидов. Кроме того, ФСГ способствует росту и развитию фолликулов в яичнике.
У мальчиков тенденция к повышению ФСГ выражена слабее. Возможно, лютеинизирующая функция гипофиза у мальчиков играет более важную роль в развитии гонад, чем у девочек [Скородок Л. М., Савченко О. Н., 1984].
Уровень ЛГ значительно повышается у детей обоего пола лишь к середине пубертата, когда гормонпродуцирующие системы гонад достаточно подготовлены к своей функции. Указанные закономерности отражены на рис. 9, 10, где представлены наши данные, полученные при исследовании уровня гонадотропных гормонов у детей в течение пубертата.
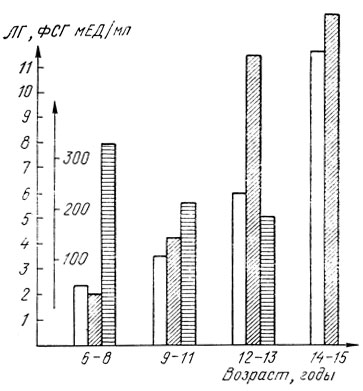
Рис. 9. Возрастная динамика средних показателей уровня гонадотропных гормонов в сыворотке крови у здоровых девочек препубертатного и пубертатного возраста. Светлые столбики - ЛГ, косо заштрихованные - ФСГ, горизонтально заштрихованные - ПР
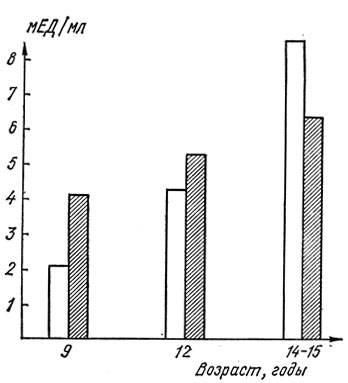
Рис. 10. Возрастная динамика средних показателей уровня гонадотропных гормонов в сыворотке крови у здоровых мальчиков препубертатного и пубертатного возраста
Кроме снижения порога чувствительности гипоталамуса к половым стероидам в системе обратной связи, в период пубертата повышается чувствительность гипофиза к стимулирующему воздействию ГТ-РГ гипоталамуса, что также приводит к повышению секреции гонадотропных гормонов. Увеличение чувствительности клеток гипофиза к стимулирующему воздействию ГТ-РГ опосредовано возрастанием уровня эндогенного ГТ-РГ [Forest М., 1976]. В последние годы преобладает мнение о ведущей роли половых стероидов, главным образом эстрогенов, в повышении чувствительности как гипоталамуса, так и гипофиза к стимулирующим воздействиям эндогенного ГТ-РГ [McCann S., 1974; Ojeda S., 1976].
В литературе много сообщений посвящено ответу гонадотропных гормонов на стимуляцию экзогенным ГТ-РГ в различные возрастные периоды [Jarnier Р., 1974; Lovrencic М., 1976; Dickerman Z. et al., 1977]. Установлено четкое повышение уровня гонадотропных гормонов в ответ на введение люлиберина уже на самых ранних этапах пубертата. Базальные значения гонадотропинов в этот период еще не менялись и отсутствовало достоверное повышение уровня периферических стероидов.
На рис. 11 и 12 представлены данные об изменении уровня гонадотропных гормонов в ответ на стимуляцию люлиберином (ЛГ-РГ 100 мкг; Relefact, ФРГ), полученные нами при обследовании детей в начальном и в зрелом пубертате (P1 и P4 по Таннеру). Результаты пробы четко указывают на зависимость гонадотропной чувствительности от возраста й пола ребенка. Препубертатный период характеризуется невысокой чувствительностью ЛГ-продуцирующих клеток к люлиберину. В пубертатный период та же самая доза ЛГ-РГ вызывает более выраженное повышение уровня ЛГ, что объясняется изменением порога чувствительности гипофиза. На протяжении всего пубертата резерв ЛГ у девочек значительно выше, чем у мальчиков. Чувствительность ФСГ-продуцирующих клеток на протяжении пубертата меньше изменяется у детей обоего пола, хотя относительный подъем более выражен на начальных стадиях.
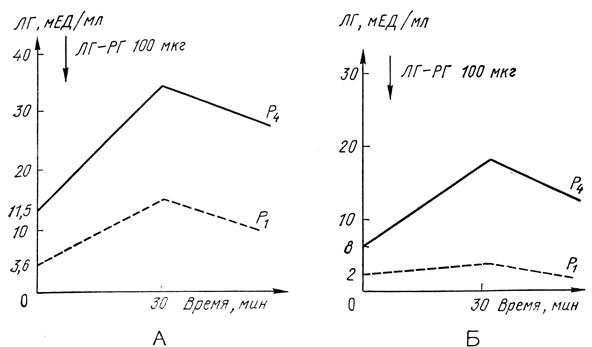
Рис. 11. Реакция лютеинизирующего гормона (ЛГ) на внутривенное введение люлиберина (ЛГ-РГ) у здоровых детей препубертатного (P1) и пубертатного возраста (Р4)
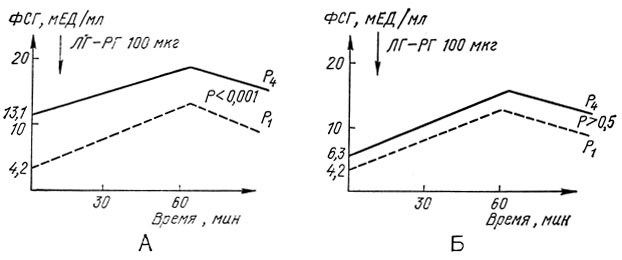
Рис. 12. Реакция фолликулостимулирующего гормона (ФСГ) на внутривенное введение люлиберина (ЛГ-РГ) у здоровых детей препубертатного (P1) и пубертатного возраста (Р4). А - У девочек; Б - у мальчиков
Пубертатная перестройка затрагивает не только изменения пороговой чувствительности гипоталамуса к ингибирующим и стимулирующим воздействиям, но и чувствительность ткани самих гонад, повышает секрецию половых гормонов в ответ на гонадотропную стимуляцию. Это объясняется главным образом увеличением количества рецепторов к гонадотропинам в гонадах. Изменяется также количество секретируемых гормонов. У мальчиков в пубертатном периоде яички секретируют тестостерон и его активный метаболит дигидротестостерон, а в допубертатном периоде преобладает секреция андростендиола и его производных [Скородок Л. М., Савченко О. Н., 1984].
По мере развития пубертата происходит дальнейшее созревание гонадостата. Повышение уровня гонадотропных гормонов не постоянное, а скачкообразное, с постепенным увеличением их общего уровня и амплитуды колебаний [Chipman J., 1980]. Чрезвычайно характерно для становления пубертата ночное увеличение выброса гонадотропинов, главным образом ЛГ [Kapen S., 1974; Kulin H. et al., 1976]. Этот показатель гонадотропной секреции является важным биологическим индексом пубертатной зрелости. За пульсирующий пубертатный выброс гонадотропинов в основном ответствен ГТ-РГ, так как имеются данные, доказывающие эпизодичность его секреции [Chipman J., 1980].
Ночным пиком ЛГ в пубертате отмечен и более высокий подъем тестостерона у мальчиков [Judd Н., 1974]. У девочек максимальная концентрация эстрадиола наблюдается в дневные часы. Это различие, видимо, обусловлено особенностями стероидогенеза в яичниках. Как известно, образование эстрадиола требует взаимодействия между овариальными клетками различных типов. Клетки theca interna синтезируют андрогены, которые поступают в гранулезные клетки, выполняющие ароматизацию стероидов под влиянием ФСГ. Длительность этого двухфазного процесса может объяснять задержку появления эстрадиола в кровяном русле после повышения уровня гонадотропинов.
По мере развития пубертата к ночным импульсам присоединяются дневные, происходит увеличение амплитуды и частоты импульсов с формированием зрелого базового так называемого цирхорального типа секреции ЛГ с частотой импульсов 60-90 в минуту.
Пубертат у девочек завершают развитие циклического центра гипоталамуса и появление положительной обратной связи с эстрогенами [Winter J., 1973; Sizonenko P., 1978]. Постепенное нарастание общего уровня гонадотропинов приводит к повышению уровня эстрадиола, который, достигая определенного критического значения, возбуждает циклический центр гипоталамуса. Оттуда стимулирующие импульсы направлены к тоническому центру, выделяющему повышенное количество ГТ-РГ. Этот центр, воздействуя на гипофиз, вызывает высокий выброс ЛГ, достаточный для овуляции. Становление овуляторного цикла завершает формирование половозрелого женского организма (рис. 13, 14).
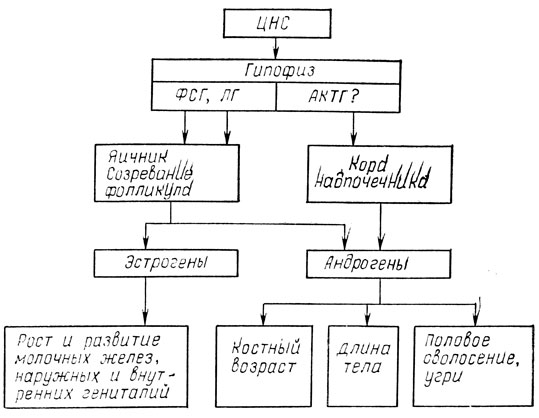
Рис. 13. Механизм эндокринных влияний в пубертатном периоде у девочек
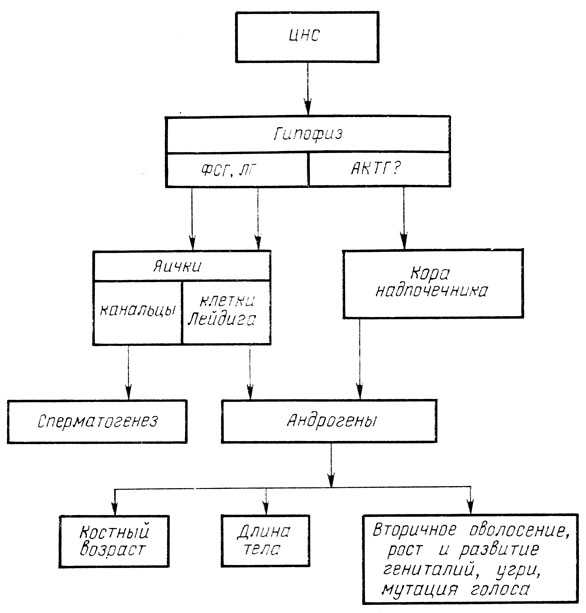
Рис. 14. Механизм эндокринных влияний в пубертатном периоде у мальчиков
Таким образом, процесс становления гонадной функции человека является длительным и сложным. Он начинается во внутриутробном периоде и завершается в конце пубертата. В основе этого процесса лежит изменение чувствительности гипоталамо-гипофизарного комплекса к существующему уровню гормонов.
До сих пор остается невыясненным вопрос о первоначальном механизме, "запускающем" пубертатное развитие, меняющем порог чувствительности гонадостата до значения, свойственного взрослому организму. Эксперименты с разрушением определенных отделов гипоталамуса и последующим преждевременным половым развитием у животных позволяют предположить, что переход к половому созреванию связан с ослаблением тормозных влияний некоторых отделов гипоталамуса на гонадотропную функцию гипофиза. Возможно, это ослабление тормозных влияний связано с падением содержания α-адренорецепторов в гипоталамусе [Wilkinson М., 1979].
Широко известно тормозящее влияние эпифиза на половое развитие. В постпубертатный период эпифиз претерпевает инволюцию. Возможно, что мелатонин и другие антигонадотропные субстраты эпифиза и являются допубертатными факторами, ингибирующими активность гипоталамуса [Sizonenko Р., 1978]. В работах R. Silman (1979) доказано значительное снижение содержания мелатонина и его производных в кровяном русле с наступлением пубертата у мальчиков. Правда, остается неясной функция мелатонина у девочек.
В последние годы накоплены факты, позволяющие предположить роль пролактина в инициации пубертата. Экспериментальные работы показали, что имплантация пролактина в срединное возвышение гипоталамуса крыс или подкожные инъекции гормона ускоряют половое созревание [Advis J. et al., 1979]. Это можно объяснить способностью пролактина увеличивать число рецепторов к ЛГ в гонадах. Кроме того, существует мнение о ведущей роли пролактина в пуске адренархе - препубертатной стадии, на которой у детей обоего пола надпочечники продуцируют много андрогенов. В настоящее время широко обсуждается стимулирующее влияние надпочечниковых андрогенов на созревание гонадостата. Это предположение подтверждается, как указывалось, случаями раннего истинного полового развития у детей с врожденной дисфункцией коры надпочечников.
Однако в ряде случаев у детей с преждевременным половым развитием церебрального генеза стадия адренархе отсутствует или выражена недостаточно, что доказывает возможность изолированного созревания гипоталамо-гипофизарно-гонадного взаимоотношения без участия адреналовых структур (рис. 15, 16).
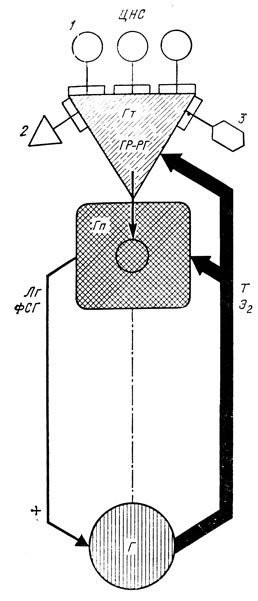
Рис. 15. Схема нейрогуморальной регуляции половой функции у ребенка допубертатного возраста. Высокая чувствительность гонадостата к ингибирующему влиянию половых стероидов. 1 - стимулирующие влияния ЦНС; 2 - ингибирующее воздействие эндогенных опиатов; 3 - ингибирующее воздействие эпифиза; Гт - гипоталамус; Гп - гипофиз; Г - гонады
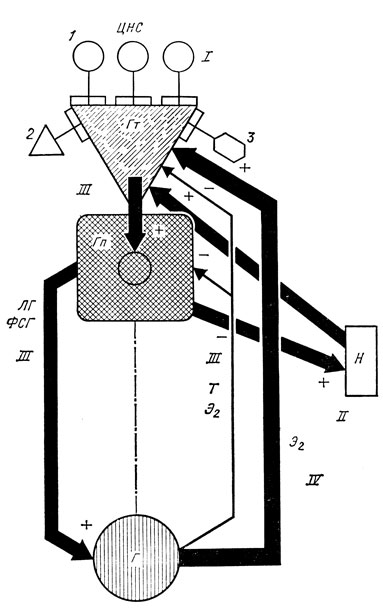
Рис. 16. Схема нейрогуморальной регуляции половой функции у ребенка в период пубертата. I - ослабление ингибирующих влияний эпифиза, эндогенных опиатов. Усиление стимулирующих влияний ЦНС; II - становление фазы адренархе: повышение секреции андрогенов надпочечников под влиянием АКТГ и других невыясненных адренотропных факторов. Воздействие надпочечниковых андрогенов на ГТ-РГ-секретирующие ядра гипоталамуса; III - снижение чувствительности гипоталамуса и гипофиза к подавляющему влиянию половых стероидов. Повышение синтеза и секреции ГТ-РГ и ЛГ и ФСГ; VI - увеличение синтеза и секреции половых стероидов; повышение чувствительности гипоталамуса к стимулирующему воздействию эстрогенов; 1 - стимулирующие влияния ЦНС; 2 - ингибирующее влияние эндогенных опиатов; 3 - ингибирующее воздействие эпифиза; Гт - гипоталамус; Гп - гипофиз; Г - гонады; Н - надпочечники
Ряд авторов придают большое значение в пубертатной перестройке гонадостата изменениям соотношений ЛГ и ФСГ [Burr J., 1970; Sizonenko P., 1970]. Известно, что ФСГ значительно увеличивает чувствительность гонад к стимулирующему воздействию ЛГ из-за возникновения новых ЛГ-рецепторов [Odell W., Swerdloff R., 1975]. Так как изменение ответа гонад на ЛГ является главным фактором в половом созревании, а сам ЛГ определяется воздействием ФСГ, ряд авторов считают, что половое созревание может наступить при одном лишь повышении уровня ФСГ [Бабичев В. Н., 1981]. Тем не менее клинические и экспериментальные данные подтверждают повышение уровня ФСГ по сравнению с уровнем ЛГ в начале пубертата, что может служить причиной пубертатной перестройки гонадостата.
Существует еще одна гипотеза о причине изменения чувствительности гипоталамуса к циркулирующим в крови половым гормонам. L. Martini (1974) доказал значительное снижение активности 5-α-редуктазы у крыс по мере их развития; 5-α-редуктазный метаболит тестостерона дигидротестостерон (ДНТ) эффективно тормозит гонадостат. К моменту половой зрелости у крыс соотношение ДНТ/Т сдвигается в сторону последнего, для торможения гипоталамо-гипофизарной системы требуются гораздо более высокие концентрации тестостерона, чем дигидротестостерона. Однако такой механизм "запуска" пубертата маловероятен, так как существенное снижение активности 5α-редуктазы отмечается только в позднем пубертате.
Таким образом, становление гонадостата является сложным, многогранным процессом, требующим сочетания многих факторов, формирующих в конечном итоге изменения чувствительности гонад к гонадотропинам в системе положительной обратной связи и чувствительности гипоталамуса к половым стероидам в системе отрицательной обратной связи.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://sohmet.ru/ 'Sohmet.ru: Библиотека по медицине'