БАЗАЛЬНЫЕ ЯДРА
Расстановка ударений: БАЗА`ЛЬНЫЕ Я`ДРА
БАЗАЛЬНЫЕ ЯДРА [позднелат. basalis относящийся к основанию; син.: центральные узлы, подкорковые ядра (nuclei subcorticales)] - скопления серого вещества в толще больших полушарий головного мозга, участвующие в коррекции программы сложного двигательного акта и формировании эмоционально-аффективных реакций.
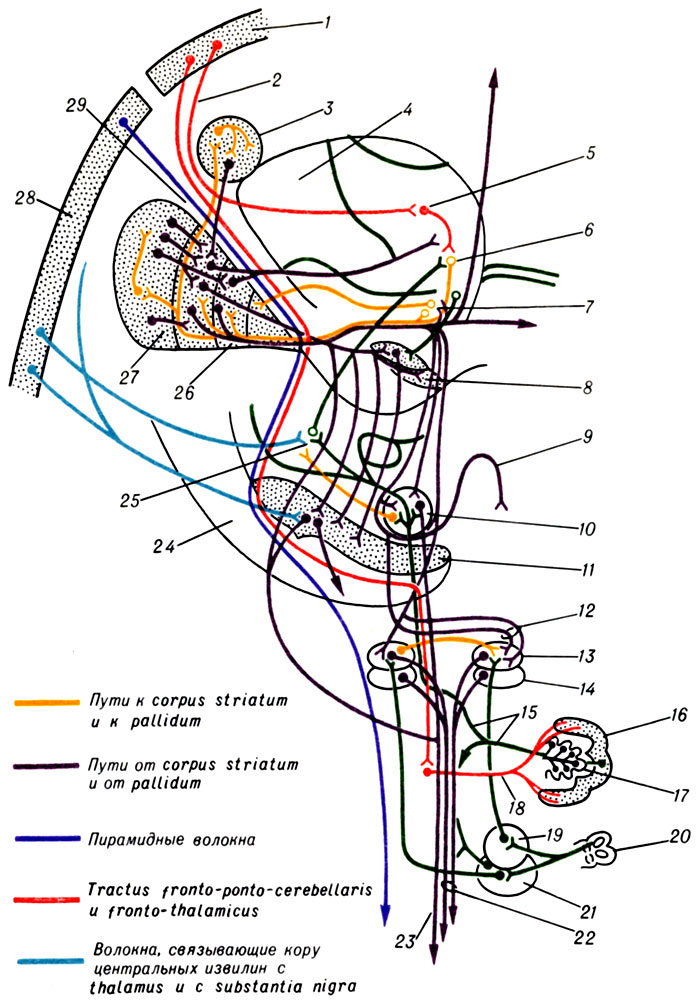
К ст. Базальные ядра. Рис. 1. Схема главных связей экстрапирамидальной системы (по С. и О. Фогт): 1 - cortex prefrontalis; 2 - tractus frontothalamicus; 3 - nucleus caudatus; 4 - thalamus; 5 - nucleus medialis thalami; 6 и 25 - nucleus ventralis thalami; 7 - nucleus campi Foreli (BNA); 8 - nucleus subthalamicus; 9 - decussatio Foreli (BNA); 10 - nucleus ruber; 11 - substantia nigra; 12 - comissura post.; 13 - nucleus Darkschewitschi; 14 - nucleus interstitialis; 15 - pedunculi cerebelli superiores (tractus cerebellotegmentalis); 16 - cerebellum; 17 - nucleus dentatus; 18 - pedunculi cerebelli medii; 19 - nucleus vestibularis sup.; 20 - canalis semicircularis; 21 - nucleus vestibularis lat.; 22 - fasciculus longitudinalis medius; 23 - fasciculus rubrospinalis; 24 - crus cerebri; 26 - globus pallidus; 27 - putamen; 28 - area gigantopyramidalis; 29 - capsula interna
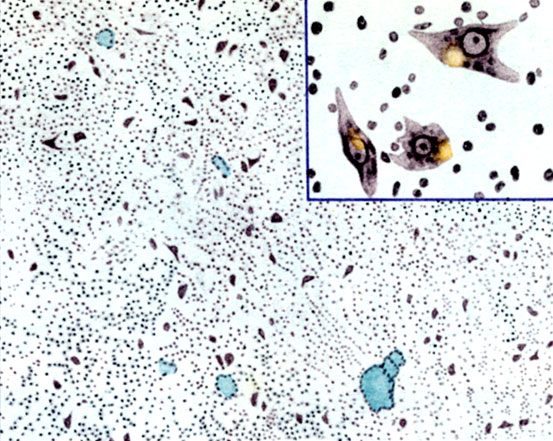
К ст. Базальные ядра. Рис. 2. Putamen. Клетки при большом и малом увеличении
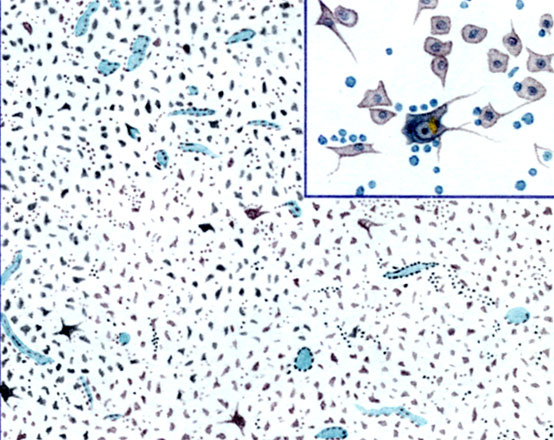
К ст. Базальные ядра. Рис. 3. Globus pallidus. Клетки при большом и малом увеличении
Первые сведения по морфологии Б. я. встречаются в работах Бурдаха (K. F. Burdach), 1819; И. П. Лебедева, 1873; Антона (Anton), 1895; Капперса (C. A. Kappers), 1908, и др. Большой вклад в изучение Б. я. внесли анатомические и клинико-морфологические исследования С. Фогт и О. Фогта (C. Vogt, O. Vogt), 1920; М. О. Гуревич, 1930; Фуа и Николеско (Foix, Nicolesco), 1925; Е. К. Сеппа, 1949; Т. А. Леонтовича, 1952, 1954; Н. П. Бехтеревой, 1963; Э. И. Канделя, 1961; Л. А. Кукуева, 1968, и др.
Б. я. наряду с расположенной по поверхности полушарий корой мозга (cortex cerebri) составляют клеточное вещество конечного мозга (telencephalon). В отличие от коры, имеющей строение экранных центров (характеризующееся определенными цитоархитектоническими признаками: четким выделением слоев, вертикальной ориентацией большинства нейронов, их дифференцировкой по форме и величине в зависимости от их положения в разных слоях), Б. я. имеют строение ядерных центров, где подобная структурная организация отсутствует. Нередко эти ядра называют подкоркой. К ним относятся: хвостатое ядро (nucleus caudatus), чечевицеобразное ядро (nucleus lentiformis, s. nucleus lenticularis), ограда (claustrum) и миндалевидное тело (corpus amygdaloideum). К Б. я. относят также базальный комплекс ядер, расположенный между передним продырявленным веществом (substantia perforata anterior) и передней частью бледного шара (globus pallidus), принадлежащий септальной области (см.).
Сравнительная анатомия. Исследования развития Б. я. в фило- и онтогенезе показали, что хвостатое ядро и скорлупа чечевицеобразного ядра (putamen) развиваются из ганглиозного бугра, расположенного на нижней стенке бокового желудочка. Они представляют единую клеточную массу, к-рая у высших позвоночных разделяется волокнами передней ножки внутренней капсулы (crus anterior capsulae internae). Ввиду общности происхождения и сохраняющегося в течение всей жизни соединения головки хвостатого ядра и переднего отдела скорлупы полосками серого вещества, чередующимися с белыми пучками волокон внутренней капсулы, хвостатое ядро и скорлупу объединяют под названием "полосатое тело" (corpus striatum), или "стриатум" (striatum). Т. к. полосатое тело является филогенетически более поздним образованием, чем медиально расположенная часть чечевицеобразного ядра - бледный шар, состоящий из наружного и внутреннего члеников, его называют "неостриатум" (neostriatum), а бледный шар - "палеостриатум" (paleostriatum). Последний в наст, время выделяют в особую морфологическую единицу под названием "паллидум" (pallidum). Исследования Л. А. Кукуева (1968) показывают, что наружный и внутренний членики бледного шара имеют различное происхождение. Наружный членик, как и скорлупа, развивается из ганглиозного бугра конечного мозга; внутренний членик - из промежуточного мозга и гомологичен энтопедункулярному ядру субприматов (расположено у них в мозге над зрительным трактом, т. е. его топография сходна с топографией внутреннего членика бледного шара на ранних стадиях развития зародыша человека). В процессе как филогенетического, так и онтогенетического развития происходит перемещение внутреннего членика по направлению к наружному, в результате чего они сближаются.
Б. я. различно представлены в мозге разных классов позвоночных животных. Так, у рыб и амфибий Б. я. представлены лишь бледным шаром, хвостатое ядро и скорлупа появляются впервые у рептилий, особенно хорошо они развиты у птиц. У млекопитающих (хищных и грызунов) бледный шар представлен единым образованием, у человека он состоит из двух члеников, разделенных прослойкой белого вещества. Размеры полосатого тела уменьшаются по мере развития головного мозга в филогенезе. Из млекопитающих у низших насекомоядных оно составляет 8% от величины всего конечного мозга, у тупайя и полуобезьян - 7%, а у обезьян - 6%.
В онтогенезе полосатое тело можно дифференцировать в начале 2-го месяца эмбрионального развития. На 3-м месяце развития головка хвостатого ядра вдается в полость бокового желудочка. Латеральнее хвостатого ядра формируется скорлупа, к-рая вначале нерезко отграничена от остальных частей полушария. Миндалевидное тело занимает особое положение среди Б. я.; на ранних стадиях эмбрионального развития оно отделяется от полосатого тела, цитологическая дифференцировка происходит в нем позднее, чем в бледном шаре, однако несколько раньше, чем в полосатом теле. Исходя из онто- и филогенетического развития, нельзя его также рассматривать и как измененную, утолщенную часть коры височной доли или как результат ее погружения внутрь и отшнуровывания. При исследовании миндалевидного тела в сравнительно-анатомическом аспекте было выявлено заметное уменьшение его размеров у млекопитающих - от низших насекомоядных, где оно вместе с палеокортексом составляет 31% от всей величины конечного мозга, до человека, в головном мозге к-рого миндалевидное тело составляет лишь 4% от всей массы конечного мозга. Исследования Развития ограды в онто- и филогенезе (И. Н. Филимонов) показали, что ее нельзя считать производным корковой пластинки или связывать по происхождению с полосатым телом. Она представляет собой промежуточное образование между этими основными клеточными массами конечного мозга.
Анатомия. Хвостатое ядро имеет грушевидную форму; его передняя часть утолщена и носит название головки хвостатого ядра (caput nuclei caudati). Она расположена в переднем отделе полушария и выступает в передний рог бокового желудочка (cornu anterius ventriculi lateralis), образуя его стенку снизу и латерально. Кзади от головки хвостатое ядро суживается и этот отдел его называется телом хвостатого ядра (corpus nuclei caudati). Тело хвостатого ядра ограничивает с латеральной стороны центральную часть бокового желудочка (pars centralis ventriculi lateralis) и описывает полукруг над зрительным бугром (thalamus) и чечевицеобразным ядром. Истонченный задний отдел хвостатого ядра, образующий часть крыши нижнего рога бокового желудочка (cornu inferius ventriculi lateralis), образует хвост хвостатого ядра (cauda nuclei caudati). Латеральная поверхность хвостатого ядра прилежит к внутренней капсуле (capsula interna), его медиальный край примыкает к конечной полоске (stria terminalis).
Чечевнцеобразное ядро имеет форму клина, основание к-рого направлено латерально, а верхушка медиально и вниз, примыкая к подбугорной области. Оно лежит латерально и несколько ниже (вентральнее) от хвостатого ядра и зрительного бугра, от к-рых отделено внутренней капсулой. Спереди и вентрально чечевицеобразное ядро соединяется с головкой хвостатого ядра тонкими полосками серого вещества. Латеральная его поверхность несколько выпуклая и располагается вертикально, гранича с наружной капсулой (capsula externa), к-рая представляет собой тонкую белую мозговую пластинку, ограниченную латерально серым веществом - оградой (claustrum). Вентральная поверхность чечевнцеобразного ядра лежит горизонтально и в средней своей части соединяется с корой в области переднего продырявленного вещества. Две тонкие мозговые пластинки, медиальная и латеральная (laminae medullares medialis et lateralis), разделяют его на три части: наружная часть, более темно окрашенная, называется скорлупой, две другие представляют собой более слабо окрашенные наружный и внутренний членики бледного шара. Ограда является узкой пластинкой серого вещества, к-рая расположена латеральнее чечевицеобразного ядра и отделена от него наружной капсулой. От коры островка (insula) ограда отделена слоем белого вещества, образующим внешнюю капсулу (capsula extrema).
Миндалевидное тело - это комплекс ядер, расположенных в области крючка парагиппокампальной извилины (uncus gyri parahippocampalis), хорошо дифференцированных и отличающихся друг от друга цитологически и цнтоархитектонически (см. Амигдалоидная область).
Гистология. Хвостатое ядро и скорлупа сходны по гистологическому строению. Серое вещество этих ядер состоит из двух видов клеточных элементов: малых и крупных клеток. Малые клетки, величиной до 15-20 мкм, имеющие короткие дендриты и тонкие аксоны, обладают нежной зернистостью и большим ядром с ядрышком. Крупные клетки, размерами до 50 мкм, в основном треугольные и полигональные, ядро их часто расположено эксцентрично, в протоплазме имеются хроматиновые зернышки и по соседству с ядром большое количество желтого липоидного пигмента. Эти клетки в норме окружены сателлитами. Отношение крупных клеток к малым в хвостатом ядре и скорлупе в среднем составляет 1:20. Как малые, так и крупные клетки имеют длинные аксоны, прослеживаемые к другим глубоким структурам мозга.
Определенные взаимоотношения между клеточными элементами и волокнами позволили Фогту (O. Vogt) указать на сходство строения полосатого тела с корой. В хвостатом ядре под эпендимой располагается зона, бедная волокнами; наружная часть этой зоны бедна ганглиозными клетками, внутренняя ими более богата. Глубже находится слой тангенциальных волокон, содержащий небольшое количество ганглиозных клеток. На основании этого Фогтом была разработана схема структурнофункциональной организации полосатого тела (цветн. табл., рис. 1): стриопетальные волокна оканчиваются на малых клетках, тесно связанных между собой и с большими клетками, от к-рых уже начинаются стриофугальные волокна. В малых клетках фибриллы не дифференцированы, в больших - распределены пучками. Миелиновых волокон в полосатом теле мало, большая часть их возникает в самом полосатом теле и служит для соединения с паллидумом; между пучками миелиновых волокон располагается густая сеть безмиелиыовых. Богатая сеть нейроглии окружает нервные клетки и нервные волокна. В паллидуме имеются клетки только очень крупные, различной формы - пирамидные, веретенообразные, мультиполярные с длинными дендритами (цветн. табл., рис. 2 и 3). В протоплазме много хроматофильных глыбок. Поверхность клеток покрыта петлеобразными концевыми тельцами - окончаниями безмиелиновых волокон, окружающих клетки и миелиновые волокна. Миелнновых волокон много больше, чем серого вещества; этим объясняется бледный цвет ядра.
Кровоснабжение Б. я. осуществляется гл. обр. от средней мозговой артерии (a. cerebri media), ветвями, идущими к полосатому телу (rr. striati). Ветви передней мозговой артерии (a. cerebri anterior) также принимают участие в кровоснабжении Б. я. Все Б. я., особенно полосатое тело, очень богаты капиллярами; распределение капилляров в полосатом теле напоминает распределение в коре; при поражениях сосудов мозга в полосатом теле особенно часто появляются очаги размягчения.
Связи базальных ядер. Афферентные волокна полосатое тело получает от зрительного бугра, от ядер нодбугорья, окружающих III желудочек, из покрышки среднего мозга (tegmentum mesencepnali) и от черного вещества (substantia nigra). Эти волокна заканчиваются около малых клеток полосатого тела, от к-рых аксоны преимущественно идут к большим клеткам, а уже от этих последних волокна идут в паллидум в составе стрио-паллидарного пучка (fasciculus striopailidalis). Волокна хвостатого ядра пересекают внутреннюю капсулу, входят в скорлупу, а затем, пронизывая мозговую пластинку, проникают в паллидум. Из скорлупы, от ее крупных клеток, волокна через мозговую пластинку также входят в паллидум. Последний является главным местом, куда направляются волокна от хвостатого ядра и скорлупы. Нек-рые авторы не отрицают возможности существования длинных волокон, идущих непосредственно из скорлупы в ствол, не прерываясь в паллидуме. Афферентные волокна, направляющиеся в паллидум, состоят из волокон, идущих: 1) непосредственно от коры; 2) из коры через зрительный бугор; 3) от полосатого тела; 4) от центрального серого вещества (substantia grisea centralis) промежуточного мозга; 5) от крыши (tectum) и покрышки (tegmentum) среднего мозга; 6) от черного вещества.
Эфферентные волокна Б. я. отходят от бледного шара. Главный пучок, выходящий из него, - лентикулярная петля (ansa lenticularis); ее волокна начинаются в хвостатом ядре, принимают участие в образовании мозговых пластинок (laminae medullares). Петля прерывается в бледном шаре. Волокна, выходящие из бледного шара, пересекают внутреннюю капсулу; на границе с ножками мозга в подбугорье они рассыпаются веерообразно и заканчиваются в переднем и латеральном ядрах зрительного бугра, в подбугорье (hypothalamus), черном веществе, подбугорном ядре (nucleus subthalamicus) и красном ядре (nucleus ruber). Часть волокон идет в составе переднего перекреста покрышки (decussatio tegmentalis anterior) на противоположную сторону, где заканчивается в одноименных образованиях. Другой пучок, выходящий из бледного шара, - лентикулярный пучок (fasciculus lenticularis). Этот пучок располагается под zona incerta, включает волокна, идущие к подбугорному ядру (вокруг к-рого образуют сумку), к зрительному бугру, красному ядру, ядру нижней оливы (nucleus olivaris), сетчатому веществу (formatio reticularis), четверохолмию [corpora quadrigemina (BNA); lamina tecti (PNA)], перивентрикулярным ядрам. Часть волокон через передний перекрест покрышки переходит на противоположную сторону и заканчивается в тех же образованиях. Описаны пути, соединяющие полосатое тело с областью воронки (infundibulum) и располагающиеся над zona incerta. От красного ядра, четверохолмия начинаются периферические экстрапирамидные волокна (tractus rubrospinalis, tractus tectospinalis). Точных данных о связи ограды и миндалевидного тела пока нет. В литературе имеются указания на связь у животных ограды с волокнами из наружной сумки, происходящими из пириформной области, на ее связь с миндалевидным телом противоположной области и вентральной областью промежуточного мозга. Установлено также, что ограда связана с корой островка. Связи миндалевидного тела - см. Амигдалоидная область.
Физиология базальных ядер. На низших ступенях эволюции (у рыб, рептилий, птиц) Б. я. являются высшими центрами координации сложного поведения. У человека и высших животных (приматов) сложную интегративную деятельность осуществляет кора больших полушарий, однако роль Б. я. не уменьшается, а лишь видоизменяется (Е. К. Сепп, 1959).
На ранних стадиях постнатального онтогенеза основная двигательная функция новорожденного - непроизвольные хаотические движения - осуществляется в основном за счет паллидума. С развитием стриатума в более поздние сроки постнатального онтогенеза отмечаются эмоциональные проявления (улыбка) и усложняются статокинетические и тонические функции (ребенок удерживает головку, выполняет содружественные движения). При рассмотрении физиологической роли Б. я. необходимо исходить из особенностей связей этих ядер с другими отделами головного мозга (Е. П. Кононова, 1959; И. Н. Филимонов, 1959; О. Затер, 1962). Б. я. характеризуются богатством афферентных и эфферентных связей с моторными зонами коры головного мозга (рис., I), с ядрами зрительного бугра (рис., II), между Б. я. (рис., III), с ядрами среднего мозга (рис., IV), а также с подбугорьем, образованиями лимбической системы и мозжечком. Важное значение для понимания физиологии Б. я. имеет учет обратных связей, идущих от них в кору головного мозга. Такой широкий спектр связей обусловливает сложность функционального значения Б. я. (объединяемых в стрио-паллидарную систему) в различных нейрофизиологических и психофизиологических процессах (В. А. Черкес, 1963; Е. Ю. Ривина, 1968; Н. П. Бехтерева, 1971). Установлено участие Б. я. в следующих нейрофизиологических функциях: а) сложные двигательные акты; б) вегетативные функции; в) безусловные рефлексы; г) сенсорные процессы; д) условнорефлекторные механизмы; е) психофизиологические процессы (эмоции). Роль Б. я. в осуществлении сложных двигательных актов заключается в том, что они обусловливают миостатические реакции, оптимальное перераспределение мышечного тонуса (благодаря модулирующим влияниям на нижележащие структуры ц. н. с., определяющие регуляцию движений).
Так, изучение функции паллидума, проведенное в условиях хронического опыта, позволило установить его важную роль в протекании сложных безусловных рефлексов различной биол. направленности - половых, пищевых, оборонительных и др.
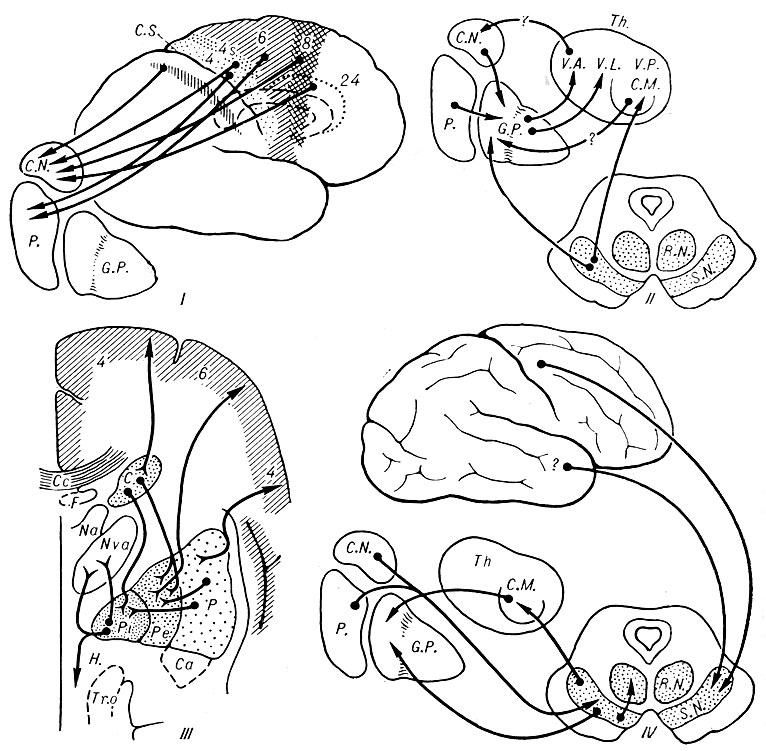
Рис. Основные афферентные и эфферентные связи (обозначены стрелками) базальных ядер с другими системами мозга (I, II, IV - по Бьюси; III - по Глису): I - связи из моторной и премоторной зон (поля 4, 4S, 6, 8, 24) коры мозга к хвостатому ядру и скорлупе; II - связи базальных ядер с ядрами зрительного бугра; III - связи между отдельными базальными ядрами и между базальными ядрами и моторной и премоторной зонами коры; IV - связи базальных ядер с черной субстанцией и красным ядром. С. N. (C - по Глису) - nucl. caudatus; V. A. (Nva - по Глису) - nucl. ventralis ant. thalami; V. L. - nucl. lateralis thalami; V. P. - nucl. ventralis post, thalami; C. M. - nucl. medialis thalami; R. N. - nucl. ruber; S. N. - substantia nigra; C. c. - corpus callosum; F - fornix; Na - nucl. ant. thalami; Tr. o. - tractus opticus; P - putamen; Pi - globus pallidus (внутренний членик); Pe - globus pallidus (наружный членик); Ca - comissura ant.; Th - thalamus; G. P. - globus pallidus; H. - hypothalamus; G. S. - sulcus centralis
Методом прямой электростимуляции паллидума показана легкость воспроизведения моторных и биоэлектрических проявлений эпилептиформ- ных реакций тонического типа. Среди важнейших функций хвостатого ядра и скорлупы следует отметить их тормозящее влияние на паллидум [Тилни и Рили (F. Tilney, H. A. Riley), 1921; Пейпс (J. W. Papez), 1942; А. М. Гринштейн, 1946, и др.]. Эффекты выключения неостриатума (полосатое тело) отражаются на функциональной активности паллидарных и среднемозговых центров (черная субстанция, ретикулярная формация ствола). Происходит их растормаживание, что сопровождается изменением мышечного тонуса и появлением гиперкинезов (см.). Многочисленные исследования влияния хвостатого ядра на условнорефлекторную деятельность и на целенаправленные движения свидетельствуют как о тормозящем, так и об облегчающем характере этих влияний, что привело к заключению о наличии двух восходящих активирующих систем: неостриарной и ретикулярной; неостриарная осуществляет влияния на кору больших полушарий как прямо, так и опосредованно, через ядра зрительного бугра. В Б. я. обнаружены явления конвергенции звуковых, зрительных, проприоцептивных импульсов. По-видимому, Б. я. являются передаточной инстанцией импульсов из ретикулярной формации в кору больших полушарий. Этим объясняются явления дезориентации, хаотической двигательной активности на фоне стимуляции хвостатого ядра и скорлупы. Важное значение полосатое тело имеет в регуляции вегетативных компонентов сложных поведенческих реакций. Раздражение неостриатума сопровождается эмоционально выразительными реакциями (мимические реакции, повышенная двигательная активность). При лечении сольных в нейрохирургических клиниках, проводимом с помощью вживленных на длительный срок электродов, показано угнетающее влияние стимуляции хвостатого ядра на выполнение интеллектуальной, речевой деятельности, на состояние памяти (Н. П. Бехтерева, 1971, и др.). Большое значение придают Б. я. в механизме развития гиперкинезов. При разрушении паллидума или его патологии наблюдается проявление гипертонии мышц, ригидность, гиперкинез. Пднако установлено, что развитие гиперкинезов есть результат выпадения функции не отдельного Б. я., а сопряжено с нарушением функций вентро-медиальных ядер зрительного бугра и центров среднего мозга, Регулирующих тонус (В. А. Черкес, 93; Н. П. Бехтерева, 1965, 1971).
Данные нейрофизиологических и клинико-неврологических исследований функций Б. я. позволяют заключить, что их физиологическое значение необходимо рассматривать в связи с другими системами мозга. Хартманн и Монаков (H. Hartmann, К. Monakow, 1960) показали, что во время сложного двигательного акта Б. я. объединяются непрерывным потоком импульсов, к-рые распространяются по определенным нейронным кругам: а) зрительный бугор - полосатое тело - зрительный бугор; б) зрительный бугор - кора полушарий - полосатое тело - бледный шар - зрительный бугор.
Функциональные взаимоотношения между Б. я. еще до конца не выяснены. Электрофизиологические исследования показали, что контроль полосатого тела над бледным шаром не является только тормозным. В острых опытах на кошках было выявлено также и облегчающее влияние хвостатого ядра на нейронную активность бледного шара, о чем свидетельствует учащение потенциалов действия отдельных элементов бледного шара под влиянием раздражения головки хвостатого ядра.
Изучение вызванных потенциалов в Б. я. показало возможность конвергенции возбуждений от различных сенсорных каналов на одном и том же нейроне [Сегундо и Макне (I. P. Segundo, X. Machne), 1956; Альб-Фессар с соавт. (D. Albe-Fessard) с соавт., 1960], и, по их мнению, ни в одной из нейронных групп Б. я. не представлена соматотопическая локализация.
Большой удельный вес афферентных морфо-функциональных связей позволяет считать, что физиологическая роль Б. я. не исчерпывается двигательной сферой. Учитывая большое значение обратных связей и тесное взаимодействие Б. я. с другими системами мозга, можно прийти к заключению, что роль Б. я. заключается в сличении различных афферентных воздействий для выполнения конечной двигательной задачи. Исходя из концепции П. К. Анохина о функциональной системе (1968), можно считать, что Б. я. участвуют в формировании афферентного синтеза, в коррекции программы сложного двигательного акта и в оценке результатов действия. Кроме того, функциональное состояние Б. я. отражается и на других функциях головного мозга, особенно при формировании эмоционально-аффективных реакций.
Методами исследования Б. я. являются: а) раздражение (электро- и химиостимуляция); б) разрушение - выключение (экстирпация, электро лизис); в) электрофизиологический метод (регистрация ЭЭГ и вызванных потенциалов); г) анализ динамики условнорефлекторной деятельности на фоне стимуляции или выключения Б. я.; д) анализ клинико-неврологических синдромов, а также психофизиологические наблюдения в клинике, проводимые в процессе лечения с помощью вживленных на длительный срок электродов (Н. П. Бехтерева, 1971). См. также Экстрапирамидная система.
Библиогр.: Анохин П. К. Биология и нейрофизиология условного рефлекса, М., 1968, библиогр.; Беритов И. С. Нервные механизмы поведения высших позвоночных животных, М., 1961, библиогр.; Бехтерева Н. П. Нейрофизиологические аспекты психической деятельности человека, Л., 1971, библиогр.; Ведяев Ф. П. Подкорковые механизмы сложных двигательных рефлексов, Л., 1965, библиогр.; Гранит Р. Электрофизиологическое исследование рецепции, пер. с англ., М., 1957, библиогр.; Коган А. Б. Электр офизиологическое исследование центральных механизмов некоторых сложных рефлексов, М., 1949, библиогр.; Рожанский Н. А. Очерки по физиологии нервной системы, Л., 1957, библиогр.; Сепп Е. К. История развития нервной системы позвоночных, М., 1959, библиогр.; Суворов Н. Ф. Центральные механизмы сосудистых нарушений, Л., 1967, библиогр.; Филимонов И. Н. Филогенез и онтогенез нервной системы, Многотомн. руководство по неврол., под ред. Н. И. Гращенкова, т. 1, кн. 1, с. 9, М., 1959; Черкес В. А. Очерки по физиологии базальных ганглиев головного мозга, Киев, 1963, библиогр.; Albe-Fessard D., Oswaldo-Cruz E. a. Rocha-Miranda C. Activites evoquees dans le noyau caude du chat en reponse a des types divers differences, Electroenceph. clin. Neurophysiol., v. 12, p. 405, 1960; Bucy P. C. The basal ganglia, the thalamus and hypothalamus, в кн.: Physiol, basis med. pract., ed. by C. H. Best, p. 144, Baltimore, 1966, bibliogr.; Clara M. Das Nervensystem des Menschen, Lpz., 1959, Bibliogr.; The diseases of the basal ganglia, ed. by T. J. Putnam a. o., Baltimore, 1942, bibliogr.
Источники:
- Большая медицинская энциклопедия. Том 2/Главный редактор академик Б. В. Петровский; издательство «Советская энциклопедия»; Москва, 1975.- 608 с. с илл., 8 л. вкл.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://sohmet.ru/ 'Sohmet.ru: Библиотека по медицине'