АРХИТЕКТОНИКА КОРЫ ГОЛОВНОГО МОЗГА
Расстановка ударений: АРХИТЕКТО`НИКА КОРЫ` ГОЛОВНО`ГО МО`ЗГА
АРХИТЕКТОНИКА КОРЫ ГОЛОВНОГО МОЗГА (полушарий большого мозга) - раздел науки о мозге, изучающий принципы общего строения и пространственных соотношений в коре головного мозга нервных клеток (цитоархитектоника), нервных волокон (миелоархитектоника), сосудов (ангио-архитектоника), межнейронных связей (синаптология) и нейроглии (глио-архитектоника). На основании архитектонических признаков вся поверхность больших полушарий головного мозга делится на структурные единицы различной значимости: основные территории, или зоны (zonae), области (regiones), подобласти (subregiones), поля (areae), подполя (subareae). А. к. г. м. дает основания для качественной и количественной оценки дифференцировки отдельных территорий и коры в целом в процессе как индивидуального, так и видового развития. Архитектоника существенно отличается от гистологии, предметом изучения к-рой является в основном тонкое строение отдельных тканевых элементов, причем топографические соотношения, имеющие для архитектоники ведущее значение, в гистологии или вообще не учитываются, или же играют подчиненную роль.
Архитектонические карты. Начальным этапом изучения местных особенностей строения коры полушарий большого мозга следует считать открытие Дженнари (F. Gennari, 1782) полоски в отделе затылочной области, к-рый в наст. время носит название area striata. Мейнерт (T. H. Meynert, 1868-1872) исследовал кору больших полушарий головного мозга микроскопически и разделил ее на пять слоев: молекулярный, наружный зернистый, пирамидный, внутренний зернистый и слой веретенообразных клеток. В это описание слоев в последующем внесен ряд уточнений; так, нижний этаж разделен Льюисом (R. Lewis, 1878) и Бродманном (K. Brodmann, 1907) наслои Y и VI (рис. 4), а Фогтом (O. Vogt) - на слои Y, VI и VII.
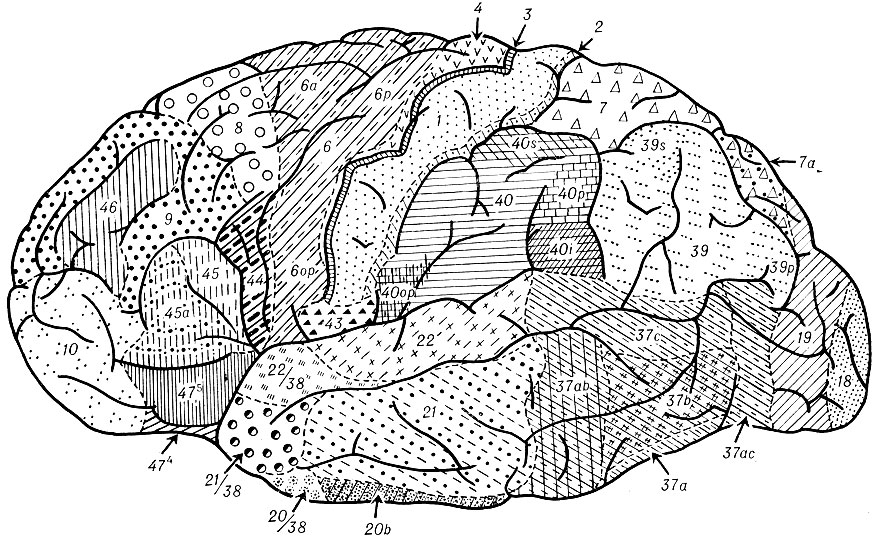
Рис. 1. Цитоархитектоническая карта коры большого мозга (Институт мозга АМН СССР). Наружная поверхность полушария. Цифрами обозначены поля новой коры, цифрами с буквами - подполя новой коры (характеристику полей и подполей см. в тексте статьи)
Большое значение имели работы киевского анатома В. А. Беца (см.), к-рый открыл гигантские пирамидные клетки в области предцентральной извилины и установил значение этой области как двигательной зоны, а также описал архитектоническое строение и ряда других областей.
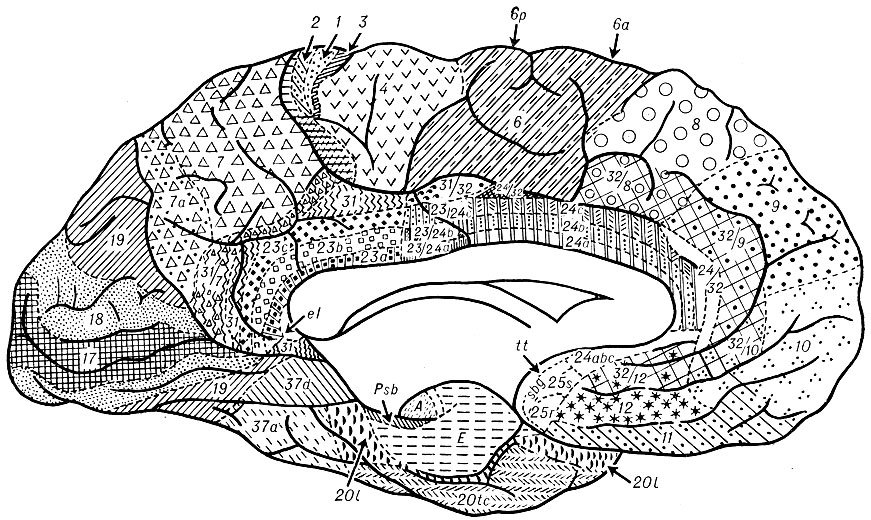
Рис. 2. Цитоархитектоническая карта коры большого мозга (Институт мозга АМН СССР). Внутренняя поверхность полушария. Цифрами обозначены поля новой коры, буквами - древней, старой и промежуточной коры. Цифрами с буквами обозначены подполя новой коры (характеристику полей и подполей см. в тексте статей)
В результате систематического изучения строения коры полушарий большого мозга появились архитектонические карты, составленные Кемпбеллом (A. Campbell, 1905), Смитом (E. Smitb, 1907), Бродманном (1903-1909), С. Фогт и О. Фогтом (C. Vogt, O. Vogt, 1919-1920), Экономо и Коскинасом (C. Economo, G. N. Koskinas, 1925), Ин-том мозга АМН СССР (1949). Карта Кемпбелла составлена на основе изучения как цито-, так и миелоархитектоиики коры полушарий большого мозга человека, орангутана, шимпанзе, собаки, кошки и свиньи. В коре полушарий большого мозга человека Кемпбелл выделил примерно 20 полей. В картах Смита деление на поля было проведено на основании только макроскопического изучения срезов коры. Бродманн на основании цитоархитектонического исследования выполнил карты мозга человека, обезьян, полуобезьян, хищных, грызунов, насекомоядных и рукокрылых. Основные границы полей коры головного мозга человека, данные Бродманном, совпадают с границами их на карте Кемпбелла, но на нарте Бродманна деление более детальное. Карта С. Фогт и О. Фогта, основывающаяся на изучении миелоархитектоиики коры полушарий большого мозга, дает деление" гораздо более дробное. Более дробное деление, чем карта Бродманна, дает и цитоархитектоническая карта Экономо и Коскинаса, к-рые делят всю поверхность полушарий большого мозга на семь долей, внося поправку, что цитоархитектонические их границы не совпадают точно с границами, принятыми анатомией.
Цитоархитектоническая карта Ин-та мозга (рис. 1 и 2) была составлена в результате сравнительноанатомического и онтогенетического исследования коры больших полушарий мозга человека. Эволюционный подход позволил выделить пять генетически обособленных территорий коры (неокортекс, архикортеке, палеокортекс, периархикортекс и перипалеокортекс), из к-рых две последние - межуточные - являются самостоятельными архитектоническими единицами, что не учитывалось во всех предыдущих цитоархитектонических картах. В цитоархитектонической карте Ин-та мозга сохраняется для обозначения полей новой коры цифровая номенклатура Бродманна: для областей древней, старой и промежуточной коры применяются буквенные обозначения. Но и в карте новой коры оказались необходимыми известные коррективы к карте Бродманна. Теменная область, единая у Бродманна, подразделяется на верхнюю и нижнюю.
В ряде случаев на основании выявления более тонкого строения различных участков коры увеличено количество обозначений для полей новой коры, но эти обозначения сохраняют связь с обозначениями Бродманна. Так, поле, обозначенное Бродманном как поле 37, рассматривается как подобласть височной области и делится на поля 37a, 37b, 37c, 37d, 37ab и 37ac.
Цитоархитектоника (neurocytoar-chitectonica corticis).
В процессе эмбрионального развития стенка конечного мозга дифференцируется, как показали классические исследования Гиса (W. His, 1904), на четыре основных слоя: 1) прилегающий непосредственно к полости желудочка материнский слой (matrix), состоящий из густого скопления нейробластов и спонгиобластов; 2) светлый промежуточный слой, содержащий мигрирующие нейробласты; 3) корковую пластинку, образующуюся из скопления мигрирующих нейробластов; 4) бедный клетками краевой слой. Но столь полно стенка конечного мозга дифференцирована только на боковой поверхности полушария, где корковая пластинка выражена четко и хорошо отделяется от промежуточного и краевого слоев.
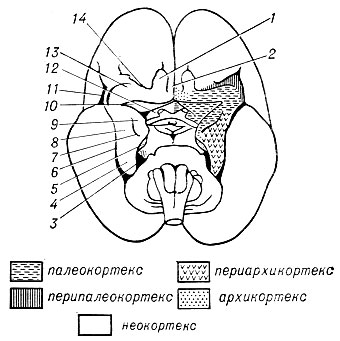
Рис. 3. Основание мозга 5-месячного плода человека: 1 - bulbus olfactorius; 2 - gyrus olfactorius medialis; 3 - sulcus parahippocampi; 4 - gyrus parahippocampalis; 5 - uncus; 6 - sulcus rhinicus posterior; 7 - sulcus semianularis; 8 - gyrus ambiens; 9 - gyrus semilunaris; 10 - tuberculum olfactorium; 11 - insula; 12 - ligamentum diagonale; 13 - gyrus olfactorius lateralis; 14 - sulcus rhinicus anterior
Это новая кора, или неокортекс (neocortex). На внутренней поверхности полушария располагается старая кора, или архикортеке (archicortex). Стенка конечного мозга отличается в этой области в ранней стадии эмбрионального развития малым количеством клеточных элементов; корковая пластинка старой коры и в дальнейшем характеризуется гораздо более слабой дифференцировкой, чем корковая пластинка неокортекса, хотя и отделена, как и там, промежуточным слоем от материнского слоя. От новой коры старая кора отделяется промежуточной периархикортикальной зоной, имеющей как в процессе онтогенетического развития, так и у взрослого явно переходный характер. На основании конечного мозга располагается древняя кора, или палеокортекс (paleocortex). В эмбриогенезе вся стенка конечного мозга в этой зоне очень широка, т. к. материнский слой образует здесь объемистые клеточные скопления в форме так наз. ганглиозных бугров, вдающихся в полость желудочков (в дальнейшем из этих бугров формируются полосатое тело, безымянное вещество и миндалевидное тело). Корковая пластинка палеокортекса на ранних стадиях не отделяется, а в постнатальном онтогенезе только слабо отделяется (в отличие от новой и старой коры) от этих клеточных скоплений, поэтому древнюю кору называют также полуотделенной корой (cortex semiseparatus). Древняя кора, так же как и старая, не соприкасается непосредственно с новой, а отделяется от нее промежуточной зоной - перипалео-кортикальной, сохраняющей, так же как и периархикортикальная зона, во всем процессе развития переходный характер.
В целом, т. о., кора полушарий большого мозга делится на пять основных зон, морфогенетически отличающихся друг от друга по своей закладке и онтогенетическому развитию: древнюю кору (палеокортекс); старую кору (архикортекс); промежуточную периархикортикальную зону и промежуточную перипалеокортикальную зону; новую кору (неокортекс) (рис. 3).
Древняя кора, или палеокортекс, имеет наименее сложное строение и характеризуется примитивной структурой корковой пластинки, слабо отделенной и у взрослого от подкорковых клеточных скоплений. В филогенезе она появляется особенно рано, в виде ольфакторной коры у селахий [Куленбек (H. Kuhlenbeck)]. В состав древней коры у животных входят обонятельный бугорок с корковой пластинкой (tuberculum olfactorium), очень слабо отделенной от головки хвостатого ядра (caput nuclei caudati), и диагональная область с корковой пластинкой, не отделяющейся от безымянного вещества, обладающего крупными клетками. Обонятельный бугорок занимает передний отдел переднего продырявленного вещества (substantia perforata ant.), диагональная область - задний его отдел и паратермина льную извилину [gyrus paraterminalis (PNA)]. В состав древней коры входит также периамигдалярная область с корковой пластинкой, слабо отделенной от миндалевидного тела (corpus amygdaloideum), располагающаяся на внутренней поверхности височной доли в ее верхне-внутреннем отделе. К древней коре относится и прозрачная перегородка (septum pellucidum) с особенно сильно редуцированной корковой пластинкой, не отделяющейся от материнского слоя, образующего ядро перегородки (nucleus septi). Наконец, самый наружный отдел древней коры у животных составляет препириформная область, корковая пластинка к-рой близко подходит к островкам серого вещества, составляющим нижний отдел ограды (claustrum). Она занимает в основном латеральную обонятельную извилину (gyrus olfactorius lat.), ограничивающую снаружи и спереди переднее продырявленное пространство.
Старая кора, или архикортеке, появляется в процессе филогенеза позднее, чем древняя кора; у амфибий она только намечена, но у рептилий выражена уже очень четко и сложно дифференцирована на области и поля. В отличие от древней коры, старая кора обладает полностью отделенной от подкорки корковой пластинкой. В то же время она резко отличается и от новой коры, т. к. остается и у взрослого однослойной или только со слабо намеченным расслоением. В состав старой коры входят располагающиеся в глубине гиппокамповой борозды гиппокамп (hippocampus) с очень крупными клетками, разделяющийся на ряд полей (subiculum, h1, h2, h3, h4, h5), и зубчатая извилина (gyrus dentatus) с корковой пластинкой, содержащей густо расположенные мелкие клетки. К старой же коре у животных относится и indusium griseum {PNA), покрывающая мозолистое тело, коленчатую извилину и медиальную обонятельную извилину [gyrus olfactorius med. (BNA)] и характеризующаяся сильно редуцированной корковой пластинкой, к-рая представляет собой продолжение кпереди корковой пластинки гиппокампа.
Промежуточная кора, появляющаяся в филогенезе одноврехменно с новой корой, т. е. у рептилий, и особенно четко дифференцированная у млекопитающих, делится на две зоны, поскольку она отделяет новую кору от древней (перипалеокортикальная зона) и от старой коры (периархикортикальная зона). Перипалеокортикальная зона, отделяющая от новой коры древнюю кору, занимает очень небольшой у человека нижний отдел островковой области, в основном представленной формациями новой коры. Периархикортикальная зона, отделяющая от новой коры старую кору, занимает парагипнокампальную извилину (gyrus parahippocampalis) и самый нижний отдел лимбической области. На парагиппокампальной извилине периархикортикальная зона делится на пресубикулярную и энторинальную области, причем последняя дифференцирована на подобласти и поля и обнаруживает очень сложное расслоение. Ее слои возникают, однако, в гораздо более ранние сроки, чем слои новой коры, и имеют иное происхождение и строение.
Новая кора, или неокортекс (син.: гомогенетическая кора Бродманна, изокортекс Фогта), по своей площади значительно превосходит у человека все остальные территории коры, вместе взятые, составляя примерно 96% всей поверхности полушарий большого мозга. Новая кора появляется впервые в процессе эволюции только у рептилий, но у них она незначительна по размерам и сравнительно проста по строению (так наз. боковая кора). Типичное многослойное строение новая кора получает только у млекопитающих, где на определенной стадии развития (5-6 мес. внутриутробной жизни) она расщепляется, как показал Бродманн, на шесть слоев, чем резко отличается от гетерогенетической коры (древняя, старая и промежуточная кора), где такого расслоения нет. Шестислойное строение гомогенетическая новая кора сохраняет и у взрослого человека (гомотипическая кора Бродманна), но в нек-рых полях число слоев или увеличивается, или уменьшается (гетеротипическая кора Бродманна).
Для основных слоев новой коры характерно следующее строение (рис. 4). Слой I, молекулярная пластинка (lamina molecularis), по своему происхождению не принадлежит корковой пластинке, а возникает на самых ранних стадиях внутриутробного развития как краевой слой стенки конечного мозга. У взрослого он очень беден клетками.
Слой II, наружная зернистая пластинка (lamina granularis externa), характеризуется, напротив, большим количеством густо расположенных клеток. Клетки относятся гл. обр. к зерновидным невроцитам (neurocytus granularis), имеющим очень небольшие размеры, кругловатую или угловатую форму. В нек-рых полях, однако, вместо зерен в слое II имеются малые пирамидальные невроциты (neurocytus pyramidalis parvus); особенно это характерно для формаций предцентральной области - "двигательная зона" (gyrus precentralis), предцентральная извилина и парацентральная долька (lobulus paracentralis) в ее переднем отделе.

Рис. 4. Цито- и миелоархитектонические слои новой коры. Основные цитоархитектонические слои обозначены римскими цифрами (подслои - цифрами с буквами); миелоархитектонические слои обозначены арабскими (подслои - цифрами с буквами)
Слой III, пирамидная пластинка (lamina pyramidalis), состоит в основном из пирамидальных невроцитов. Величина этих клеток большей частью возрастает в направлении вглубь, так что слой III в большинстве полей распадается на три подслоя: подслой III1 с малыми пирамидальными невроцитами (neurocytus pyramidalis parvus), подслой III2 со средними пирамидальными невроцитами (neurocytus pyramidalis medius) и подслой III3, содержащий большие пирамидальные невроциты (neurocytus pyramidalis magnus). Однако в нек-рых полях слой III не делится на подслои (поле 17 затылочной области), в нек-рых делится на два подслоя, в нек-рых на четыре. Также сильно меняется от поля к полю и величина клеток слоя III. В нек-рых полях (напр., в поле 17) они вообще имеют небольшие размеры, в других могут достигать очень большой величины, напр., в поле 18 на границе с полем 17, где клетки подслоя III3 приобретают местами "гигантский" характер. Сильно изменчива и густота распределения пирамидальных клеток, а также характер их расположения в вертикальном направлении: в одних полях они располагаются радиальными тяжами, в других - более или менее диффузно.
Слой IV, внутренняя зернистая пластинка (lamina granularis interna), состоит из густо расположенных малых зерновидных невроцитов круглой и угловатой формы (neurocytus granularis parvus).
Этот слой особенно изменчив и, являясь хорошо выраженными более или менее однородным в гомотипических полях, в гетеротипических полях может или отсутствовать, или же подразделяться на подслои. Пример гетеротипической коры с исчезновением слоя IV (агранулярный тип) представляют поля 4 и 6 предцентральной области (ядерная зона двигательного анализатора). От такого агранулярного типа к хорошо выраженному гранулярному типу существует ряд переходов - есть поля, где слой IV только намечен, и есть поля, где он, хотя и существует как обособленный слой, очень узок. Классическим примером гетеротипической коры с расщеплением слоя IV является поле 17 затылочной области, представляющее собой ядерную зону зрительного анализатора. Слой IV распадается здесь очень четко на подслои IVa, IVb, IVc.
Слой V, ганглионарная пластинка (lamina ganglionaris), гораздо менее густой, чем расположенный над ним слой IV, и несколько менее густой, чем находящийся под ним слой VI, состоит в основном из пирамидальных невроцитов (neurocytus pyramidalis), среди к-рых встречаются и очень большие клетки - гигантские пирамидальные невроциты - клетки Беца (neurocytus gigantopyramidalis). Он может быть единым, но большей частью делится на два подслоя. В поле 4 предцентральной области слой V распадается на три подслоя, в среднем из к-рых располагаются гигантские клетки Беца, достигающие значительной величины (до 120 мкм).
Слой VI, полиморфная пластинка (lamina multiformis), делится на два подслоя - триангулярный, состоящий преимущественно из треугольных невроцитов (neurocytus triangularis), и веретеновидный, состоящий из веретеновидных невроцитов (neurocytus fusiformis). О. Фогт выделяет эти подслои как самостоятельные слои: слой VI (lamina triangularis) и слой VII (lamina fusiformis). Строение их различно в разных полях. Особенно своеобразен колонкообразный тип строения, при к-ром клетки слоя VI располагаются колонками (поля затылочной области, особенно поле 17). Слой VI или резко отделяется от белого вещества, или же переходит в него постепенно, без резкой границы.
В соответствии с особенностями строения как всего поперечника корковой пластинки, так и отдельных ее слоев новая кора делится на ряд областей, каждая из к-рых подразделяется в свою очередь на ряд полей.
Затылочная область (поля 17, 18 и 19) характеризуется в целом небольшой величиной клеток и густым их расположением, очень светлым слоем V, колонкообразным слоем VI, преобладанием по ширине верхнего этажа поперечника коры (слои II, III и IV) над нижним этажом (слои V и VI). Центральный отдел затылочной области образует поле 17 (area striata), занимающее шпорную борозду (gyrus calcarinus) и прилегающие к ней части клина (cuneus) и медиальной височно-затылочной извилины (gyrus temporooccipitalis medialis). Мелкоклеточность и густоклеточность выражены в поле 17 особенно резко (так наз. кониокортекс, или пылевидная кора). Весьма характерными являются описанное выше расщепление слоя IV на подслои и особенно резко выраженный колонкообразный рисунок слоя VI. Окружающие гетеротипическое поле 17 кольцеобразно поля 18 и 19 относятся к гомотипической коре и по своему строению составляют как бы переход от поля 17 к полям теменной и височной областей, т. е. к полям 7, 39 и 37.
Верхняя (поля 5 и 7 с подполями 7s, 7α и 7γ) и нижняя (поле 39c подполями 39s и 39p и поле 40c подполями 40s, 40op, 40i и 40p) теменные области относятся к гомотипической коре и характеризуются четко выраженным делением поперечника коры на шесть слоев с хорошо обособленным наружным и внутренним зернистыми слоями. Характерны также радиальное распределение клеток слоя III, значительная густоклеточность и небольшая в целом величина клеток, а также постепенное, в направлении к затылочной области, уменьшение преобладания по ширине нижнего этажа коры над верхним. Поле 5, составляющее передний отдел верхней теменной области, характеризуется наряду с указанными признаками наличием очень больших клеток в слое V, напоминающих пирамидальные невроциты (гигантские клетки Беца) в поле 4 предцентральной области. Поле 7, не имеющее таких клеток, отличается, как и поле 5, от полей 39 и 40 нижней теменной области большим размером клеток и менее густым их расположением.
Постцентральная область (поля 3/4, 3, 1,2 и 43) занимает дно и заднюю стенку центральной борозды (sulcus centralis), а также поверхность постцентральной извилины (gyrus postcentralis) и заднего отдела парацентральной дольки. Дно центральной борозды занято полем 3/4, к-рое представляет по своему строению переход от постцентральной области к предцентральной области и характеризуется одновременно наличием и внутреннего зернистого слоя (признак постцентральной области), и гигантопирамидальных невроцитов (клеток Беца) в слое V (признак поля 4 предцентральной области). На задней стенке центральной борозды располагается поле 3, характеризующееся особенно малой величиной клеток и обилием малых зерновидных невроцитов, очень большой густоклеточностыо, очень светлым слоем V. В верхней части задней стенки центральной борозды и на поверхности переднего отдела постцент- ральной извилины располагается поле 1, характеризующееся очень четким разделением на шесть слоев и резко отличающееся от поля 3 значительной величиной клеток слоев III, V и VI. Примыкающее сзади к полю 1 поле 2 отличается от него только небольшими деталями в строении. Поле 43, занимающее operculum, является комплексом структур, из к-рых лишь центральные отделы имеют типичные признаки данной области.
Предцентральная область (поля 4 и 6 с подполями 6a, 6p и 6op) располагается на поверхности предцентральной извилины и переднего отдела парацентральной дольки, на передней стенке центральной борозды и на поверхности заднего отдела верхней и средней лобных извилин (gyrus frontalis sup. et med.) и характеризуется прежде всего тем, что слой IV не выражен, а слой II выражен слабо или отсутствует. Характерными признаками являются также слабая отделенность слоев друг от друга (слабая горизонтальная исчерченность), наличие особенно хорошо сформированных пирамидальных невроцитов, небольшая густоклеточность, большая ширина как всей корковой пластинки, так и особенно нижнего ее этажа (слои V и VI). Поле 4 (area gigantopyramidalis) характеризуется еще наличием гигантопирамИдальных невроцитов в слое V, к-рые придают этому полю чрезвычайно типический вид. Поле 6 отличается от поля 4 в основном только отсутствием гигантских клеток Беца.
Лобная область включает в свой состав многие поля [8, 9, 46, 10, 11, 12, 44, 45, 47 (с подполями 471472, 473, 474, 475), 32 (с подполями 32/8, 32/9, 32/10, 32/12)], из к-рых самое заднее поле 8 еще напоминает по строению предцентральную область, т. к. слой IV едва намечен. Однако, в отличие от располагающегося далее кзади поля 6, в поле 8 этот слой все же имеется. В остальном поля лобной области представлены гомотипической корой с очень ясным делением на шесть основных слоев.
Височная область (верхняя височная подобласть с полями 41, 41/42, 42, 22, 22/38 и 52, средняя височная подобласть с полями 21 и 21/38, нижняя височная подобласть с полями 20tc, 201, 20b и 20/38 и височно-теменно-затылочная подобласть с полями 37a, 37b, 37c, 37d, 37ab и 37ac) занимает всю височную долю до ринальной борозды (sulcus rhinalis), отделяющей от нее парагиппокампальную извилину, покрытую промежуточной корой, отграничивающей новую кору от старой (энторинальная и пресубикулярная области, E и Psb). Поля височной области имеют почти полностью гомотипическое строение, хорошо выраженные зернистые слои и большей частью хорошо выраженную радиальную исчерченность, особенно типическую в нек-рых полях в слое IV, к-рый распадается на своеобразные колонки. Особое строение имеет кора в задней части верхней поверхности височной доли, скрытой в латеральной (сильвиевой) борозде (поля 41 и 42). В поле 42, расположенном ближе к выходу на свободную поверхность, обращают на себя внимание наряду с мелкими густо расположенными клетками сравнительно большие клетки в подслое III. Поле 41, расположенное в глубине латеральной борозды, имеет особенно густоклеточное строение и принадлежит, как и поле 17 затылочной области и поле 3 постцентральной области, к пылевидной коре, или кониокортексу. К полям 41 и 42 афферентные волокна медиального коленчатого тела (corpus geniculaturn med.) подходят особенно густыми пучками, также густыми пучками к полю 17 подходят афферентные волокна из латерального коленчатого тела (corpus geniculatum lat.). Это ядерные зоны слухового и зрительного анализаторов.
Лимбическая область, занимающая поясную и парагиппокампальную извилины, в нижнем своем отделе (глубина борозды мозолистого тела) представлена промежуточной корой (перптектальная область), отделяющей taenia tecta (старая кора) от новой коры, в основном же она занята полями новой коры (ноля 23, 23/24, 24, 25, 31, 31/32 и 24/32). В заднем своем отделе лимбическая область имеет шестислойное строение, в переднем отделе она, как и предцентральная область, занята агранулярной корой (отсутствие ясно выраженных и обособленных слоев II и особенно IV).
Островковая область [островок (insula)] лишь в небольшом нижнем своем отделе представлена промежуточной корой, отделяющей новую кору от древней коры. В основном же островковая область, полностью скрытая в глубине латеральной (сильвиевой) борозды и образующая ее дно, относится к новой коре (поля 13 и 14). Задний ее отдел принадлежит к гомотинической коре и характеризуется хорошо выраженными зернистыми слоями, передний же отдел имеет, как и поля предцентральной области и переднего отдела лимбической области, агранулярное строение.
Величина поверхности основных зон и неокортикальных областей мозга человека по отношению к поверхности полушария выражается следующими цифрами в процентах (по данным Института мозга).
| Основные зоны | |
| Падеокортекс (древняя кора) | 0,6 |
| Перипалеокортекс | 0,3 |
| Архикортекс (старая кора) | 2,2 |
| Периархикортекс | 1,3 |
| Неокортекс (новая кора) | 95,6 |
| Неокортикальныеобласти | |
| Затылочная | 12,0 |
| Нижняя теменная | 7,7 |
| Верхняя теменная | 8,4 |
| Постцентральная | 5,4 |
| Предцентральная | 9,3 |
| Лобная | 23,5 |
| Височная | 23,5 |
| Островковая | 1,8 |
| Лимбическая | 4,0 |
Функциональная и биологическая значимость архитектонических характеристик коры полушарий большого мозга может быть понята гл. обр. при эволюционном подходе к проблеме изучения структурнофункциональной специфики мозга человека.
| Представители млекопитающих | Новая кора | Древняя кора | Показатель максимальных девиаций (в абс. ед.) |
|---|---|---|---|
| в % ко всей поверхности полушария большого мозга | |||
| Человек | 95,9 | 0,6 | 159,8 |
| Шимпанзе | 93,3 | 1,3 | 71,7 |
| Макак | 93,0 | 1,4 | 66,4 |
| Игрунка | 85,3 | 2,8 | 30,4 |
| Дельфин | 97,8 | 0,9 | 108,7 |
| Тюлень | 94,4 | 1,8 | 52,4 |
| Собака | 84,2 | 6,8 | 12,4 |
| Кенгуру | 69,2 | 9,3 | 7,4 |
| Муравьед | 57,2 | 19,3 | 3,0 |
| Кролик | 56,0 | 14,0 | 4,0 |
| Еж | 32,4 | 29,8 | 1,1 |
При исследовании мозга в восходящем ряду наземных млекопитающих обнаружено, что увеличение поверхности полушарии, усложнение рисунка борозд и извилин соответствует усложнению структурной организации корковой пластинки. Изучение водных млекопитающих показало значительное развитие больших полушарий при весьма невысокой организации неокортекса, что может свидетельствовать об определенной независимости эволюции признаков архитектонической дифференцировки коры от величины поверхности полушарий. Сопоставление данных площади поверхности полушарий (древняя, старая, промежуточная и новая кора в сумме) и веса головного мозга различных представителей млекопитающих (еж, кенгуру, муравьед, собака, свинья, тигр, тюлень, макак, павиан, орангутанг, шимпанзе, дельфин обыкновенный, дельфин афалина, кит финвал, человек) показало прямую пропорциональную зависимость площади поверхности полушарий большого мозга от веса головного мозга. Соотношения поверхности полушарий и веса мозга и у ежа, и у дельфина, и у человека одни и те же, несмотря на отсутствие борозд у ежа и их обилие у дельфина и человека. Процент площади коры, расположенной внутри борозд (интрасулькальный компонент), казавшийся ранее эволюционно значимым, оказался также зависящим от веса мозга (у кита - 88,1, у дельфина афалина - 85,0; у дельфина обыкновенного - 78,6 и у человека всего лишь 64,8).

Таблица 2. Поверхность областей новой коры человека и некоторых видов обезьян в % к поверхности коры головного мозга всего полушария большого мозга (по данным Института мозга АМН СССР)
Подобные соотношения (прямо пропорциональная зависимость площади поверхности полушарий от веса головного мозга) четко прослеживаются и в онтогенезе корковых формаций мозга человека. Следовательно, ни величина поверхности полушарий, ни количество борозд и извилин, ни интрасулькальный компонент сами по себе еще не могут служить признаками высокой эволюционной организации мозга.
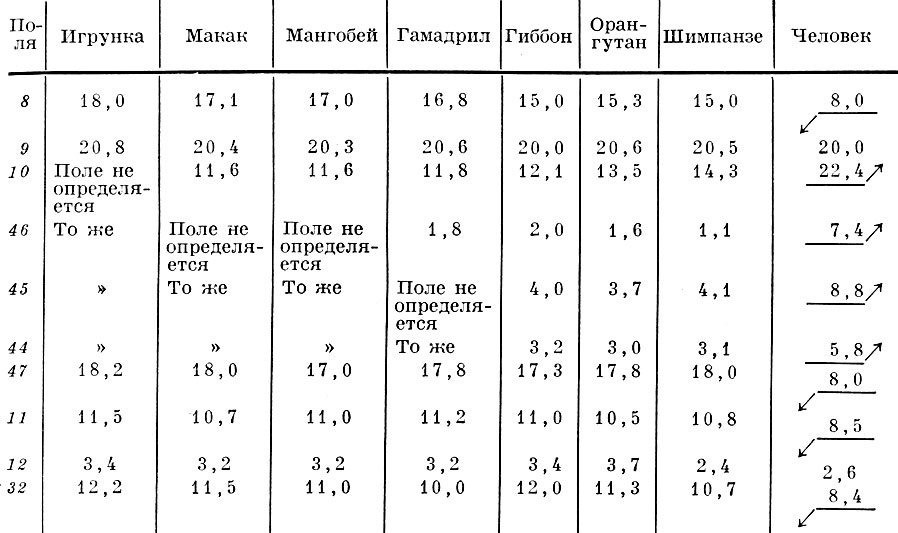
Таблица 3. Поверхность полей коры лобной области человека и некоторых видов обезьян в % к поверхности коры всей лобной области (по данным Е. П. Кононовой, Институт мозга АМН СССР)
Процесс эволюционных изменений корковых территорий больших полушарий головного мозга от низших млекопитающих к высшим и человеку заключается в тенденции дивергентного ("разнонаправленного") развития различных по генезу зон. Так, новая кора по относительной величине ко всей коре значительно увеличивается, а древняя кора явно уменьшается. II если по относительной величине новой коры человек почти не выделяется из отряда приматов и даже уступает дельфину, то по соотношению абсолютной площади новой и древней коры, как показателю максимальных девиаций (ПМД), значительно превосходит и приматов, и китообразных. Тенденция дивергентного развития филогенетически новейших и филогенетически древнейших структур в рамках генетически обособленных территорий мозга, свойственная всем наземным млекопитающим, максимального выражения достигает в мозге человека (табл. 1).
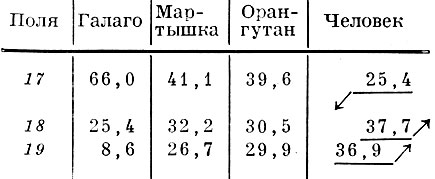
Таблица 4. Поверхность полей коры затылочной области человека и некоторых видов обезьян в % ко всей поверхности коры затылочной области (по данным Института мозга АМН СССР)
В новой коре в восходящем ряду приматов явно увеличиваются области, имеющие отношение к высшим ассоциативным и интегративным механизмам (лобная, височная, нижняя теменная), относительно уменынается филогенетически древняя часть новой коры (лимбическая область), также уменьшается проекционная зона зрительного анализатора (затылочная область) и остается стабильной во всем ряду приматов, включая человека, иредцентральная область, связанная гл. обр. с моторными функциями (табл. 2).

Таблица 5. Поверхность полей коры предцентральной области у человека и некоторых видов обезьян в % к поверхности коры всей предцентральной области (по данным Института мозга АМН СССР)
Аналогичная перегруппировка различных по генезу полей в восходящем ряду приматов происходит в пределах отдельных областей, явно прогрессирующих (табл. 3), относительно уменьшающихся (табл. 4) и относительно стабильных (табл. 5).
| Человек | ПМД | Обезьяна | ПМД |
|---|---|---|---|
| 2 лунных мес. | 4,5 | 1,5 лунных мес. | 4,6 |
| 6 лунных мес. | 40,1 | 3 лунных мес. | 42,5 |
| Новорожденный | 155,6 | Новорожденная | 64,1 |
| Взрослый | 159,8 | Взрослая | 66,4 |
Количественный анализ структурных изменений мозга в эволюции млекопитающих выявляет специфику мозга человека, заключающуюся в максимальной выраженности филогенетически новейших и минимальной - филогенетически древнейших образований в рамках генетически обособленных территорий. Сравнительное изучение онтогенеза корковых формаций мозга человека и обезьяны показало значительное нарастание поверхности полушарий мозга человека в постнатальном онтогенезе (площадь поверхности полушарий мозга новорожденного составляет ок. 20% площади поверхности полушарий мозга взрослого человека), в отличие от обезьяны, у плода к-рой к моменту рождения площадь поверхности полушарий составляет ок. 80% площади поверхности взрослой особи (И. А. Станкевич, 1964).
Кроме того, изменения ПМД (табл. 6) у плода человека и плода обезьяны в первой половине беременности имеют общий для приматов характер, тогда как перед самым рождением резкий скачок ПМД мозга человека соответствует появлению специфически "человеческих" признаков дифференцировки неокортекса (подполей).
Миелоархитектоника (myeloarchiteetonica corticis).
Миелоархитектоника особенно детально разработана исследованиями С. Фогт и О. Фогта. Миелоархитектонически новая кора распадается на шесть основных слоев, соответствующих цитоархитектоническнм слоям (рис. 4). Слой 1 называется в миелоархитектоннке тангенциальным (lamina tangentialis), т. к. в нем проходят направленные горизонтально (параллельно поверхности) тангенциальные волокна, осуществляющие внутрикорковые связи. Большей частью он состоит из четырех подслоев, но может состоять и из трех или двух подслоев. Слой 2 называется дисфиброзным (lamina disfibrosa), т. к. он беден миелиновыми волокнами. Слой 3 (lamina suprastriata) соответствует пирамидному слою в цитоархитектонической картине и, как и там, большей частью разделяется на три подслоя. В нек-рых полях в верхнем подслое выделяется описанная В. М. Бехтеревым полоска (stria laminae granularis externae), состоящая из густого скопления горизонтальных миелиновых волокон и являющаяся важным опознавательным признаком этих полей. Слой 4 характеризуется, как и полоска наружной зернистой пластинки, густым скоплением горизонтальных миелиновых волокон и называется полоской внутренней зернистой пластинки - наружная полоска Байярже (stria laminae granularis internae). Слой 5 состоит из двух подслоев: подслой 5a обозначается как lamina intrastriata и большей частью содержит гораздо меньше горизонтальных волокон, чем граничащие с ним слой 4 и подслой 5b; подслой 5b носит название полоски ганглионарной пластинки - внутренняя полоска Байярже (stria laminae ganglionaris). Слой 6 состоит из четырех подслоев, более или менее четко отличающихся друг от друга. О. Фогт выделяет нижние его подслои как слой 7.
Для разделения коры на поля большое значение имеют особенности в строении тангенциального слоя (четырех-, трех- или двухзональный тип в зависимости от количества подслоев) и в строении и соотношении полосок Байярже. Наиболее типично двухполосное строение - typus bistriatus (обе полоски хорошо обособлены), но возможны и typus unitostriatus (обе полоски слиты в одно целое вследствие большого количества волокон в располагающемся между ними подслое 5a), typus unistriatus (обособлена только полоска внутренней зернистой пластинки, а полоска ганглионарной пластинки сливается с подлежащим слоем 6) и typus astriatus (в одно целое слиты слои 4, 5 и 6 вследствие густоты обычно более светлых промежуточных прослоек, образуемых подслоем 5a и наружным подслоем слоя 6).
Наряду с горизонтально расположенными миелиновыми волокнами на миелоархитектонических препаратах в коре выступают и радиально расположенные волокна (neurofibrae radiales), к-рые входят в кору из подлежащего белого вещества и располагаются в ней лучеобразно. Особенно большое значение для миелоархитектонической дифференцировки коры имеет отношение радиальных волокон к слоям коры. Выделяют три основных типа этих связей: тип первый (typus euradiatus), где радиальные волокна достигают слоя 3, соответствует новой коре, кроме переднего отдела верхней лимбической коры; тип второй (typus infraradiatus), где радиальные волокна оканчиваются уже на границе между обоими подслоями слоя 5, занимает переднюю часть верхней лимбической извилины и также соответствует определенному отделу новой коры; тип третий (typus supraradiatus), где радиальные волокна проходят в большом количестве в слои 3 и 2 , а отсюда частично проникают и в слой 1, соответствует в целом древней, старой и промежуточной коре. Типы первый и второй О. Фогт обозначает как изокортекс, а третий тип - как аллокортекс.
Изучение архитектоники волокон коры полушарий большого мозга человека в последние годы получило большое развитие в связи с модификацией методов импрегнации (метод А. П. Авцына в модификации лаборатории архитектоники Ин-та мозга) и применением количественного анализа волокнистых структур с помощью денситометрии [Хопф (A. Hopf), 1968]. Благодаря этому выявлены не только миелиновые, но и безмиелиновые волокна, наиболее мощно представленные в верхних слоях коры (1, 2, 3, 4). Данные Ин-та мозга (1972), не противоречащие основным принципам миелоархитектонической классификации С. Фогт и О. Фогта, вместе с тем показали значительно большую вариабельность распределения радиальных и горизонтальных волокон в различных отделах коры мозга человека.
Ангиоархитектоника (angioarchitectonica corticis).
Пфейфер (R. Pfeifer, 1928) показал, что как отдельные слои коры, так и отдельные ее формации отличаются особенностями строения капиллярной сети, и дал описание ангиоархитектонического строения затылочной области (поля 17 и 18), предцентральной извилины, верхней теменной дольки (lobulus parietalis sup.), парагиппокампальной извилины. Ангиоархитектонические исследования значительно расширяют знания о строении коры больших полушарий головного мозга, достигнутые цито- и миелоархитектоникой. См. также Головной мозг (кровоснабжение).
Синаптология еще не достигла той стадии развития, когда на ее основании, как на основании данных цито- и миелоархитектоники, можно было бы выделить формации коры больших полушарий головного мозга. Однако и в этой области получены весьма важные данные. Особенно большое значение для учения о нейронных структурах имеют классические исследования Кахаля (S. Ramon у Cajal, 1909-1911), а в отечественной литературе - работы С. А. Суханова (1899). В дальнейшем исследования Лоренте де Но (R. Lorente de Nó), С. А. Саркисова, А. Д. Зурабашвили, Г. И. Полякова и многих других позволили подойти к проблеме классификации нейронов и построению принципиальных схем организации различных нейронных комплексов в корковых и подкорковых образованиях мозга.
Значительный качественный скачок в развитии синаптологии в последние десятилетия связан с применением электронной микроскопии для изучения синапса (см.).
Мейнерт (T. Meynert, 1868) сформулировал на основании своих архитектонических исследований учение о коре полушарий большого мозга как о совокупности органов, т. е. образований с различными функциями, и об архитектонике как об "органологии". В. А. Бец указывал, что деление мозга на области может быть правильно проведено только на основании тонкого изучения строения коры. Выявляя с непреложной очевидностью, что кора большого мозга дифференцирована на различно построенные формации, данные архитектонических исследований полностью опровергают представления ряда авторов об однородности ее функций во всех областях. В то же время данные архитектоники, особенно данные эволюционной морфологии, показывают, что все области коры, включая и самые примитивные, являются образованиями функционально многозначными и что нет таких областей коры, к-рые представляли бы собой "центры", обеспечивающие только одну какую-либо функцию (см. Кора головного мозга). Положение о локализуемости кортикальных функций от этого не утрачивает своего значения. Если в реализации той или иной функции и принимают участие многие архитектонические формации, то это никоим образом не значит, что участие это для всех формаций одинаково; несомненно, что деятельность нек-рых из них является при этом доминирующей, а деятельность других имеет только более или менее второстепенное значение.
Сформулированный И. П. Павловым и А. А. Ухтомским принцип локализации корковых функций предполагает динамические системы, элементы к-рых сохраняют строгую дифференцированность и играют высокоспециализированную роль в единой деятельности.
Особенно плодотворным для решения вопросов локализации функций (см.) оказалось комплексное исследование (архитектоническое и физиологическое), проведенное С. Фогт и О. Фогтом на мартышках. На границах участков, дававших на раздражение различную двигательную реакцию, проводились надрезы еще на живом мозге с целью выяснения вопроса об анатомическом субстрате этих физиологических границ.
Электрофизиологическое исследование проекционной корковой зоны зрительного анализатора показало наличие различных типов нейронов - с простыми и сложными рецептивными полями [Юнг (R. Jung), 1961; Хьюбел (D. H. Hubei) и Визел (T. N. Wiesel), 1963], причем вертикальная организация нейронных ансамблей с идентичными рецептивными полями позволила авторам выдвинуть принцип организации анализаторных корковых зон в "функциональные колонки". Морфологически данный признак описывался ранее как вертикальная исчерченность, выявляющаяся в нек-рых областях новой коры.
При изучении структурно-функциональной организации коры необходимо иметь в виду, что в осуществлении как сенсо-моторных, так и вегетативных компонентов целостного поведенческого акта принимают участие, помимо коры, подкорковые, стволовые и спинальные образования ц. н. с., взаимодействие и взаимовлияние к-рых определяет адекватность любого адаптивного процесса.
См. также Головной мозг, Кора головного моза, Патоархитектоника.
Библиогр.: Архитектоника волокон коры большого мозга человека, под ред. С. А. Саркисова, М., 1972, библиогр.; Блинков С. М. и Глезер И. И. Мозг человека в цифрах и таблицах, Л., 1964, библиогр.; Лурия А. Р. Высшие корковые функции человека и их нарушения при локальных поражениях мозга, с. 37, М., 1969, библиогр.; Развитие мозга ребенка, под ред. С. А. Саркисова, Л., 1965; Станкевич И. А. О специализации хода онтогенеза большого мозга человека, Усп. совр. биол.,т. 58, в. 3, с. 409, 1964; Филимонов И. Н. Сравнительная анатомия большого мозга рептилий, М., 1963, библиогр.; Шевченко Ю. Г. Эволюция коры мозга приматов и человека, М., 1971, библиогр.; Юнг Р. Интеграция в нейронах зрительной коры и ее значение для зрительной информации, в кн.: Теория связи в сенсорн. сист., пер. с англ., под ред. Г. Д. Смирнова, с. 375, М., 1964, библиогр.; Contemporary research methods in neuroanatomy, ed. by W. J. H. Nauta a. S. O. E. Ebbesson, N. Y. a. o., 1970, bibliogr.; Hopf A. Registration of the rnyeloarchitecture of the human frontal lobe with an extinction method, J. Hirnforsch., Bd 10, S. 259, 1968, Bibliogr.; Hubei D. H. a. Wiesel T. N. Receptive fields, binocular interaction and functional architecture in the cat's visual cortex, J. Physiol. (Lond.), v. 160, p. 106, 1962, bibliogr.; Pfeifer R. A. Die Angioarchitektonik der Grosshirnrinde, B., 1928, Bibliogr.
Источники:
- Большая медицинская энциклопедия. Том 2/Главный редактор академик Б. В. Петровский; издательство «Советская энциклопедия»; Москва, 1975.- 608 с. с илл., 8 л. вкл.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://sohmet.ru/ 'Sohmet.ru: Библиотека по медицине'