АНАЭРОБЫ
Расстановка ударений: АНАЭРО`БЫ
АНАЭРОБЫ (греч. отрицательная приставка an-, aēr — воздух и bios — жизнь) — бактерии, не требующие для своего существования и размножения свободного кислорода.
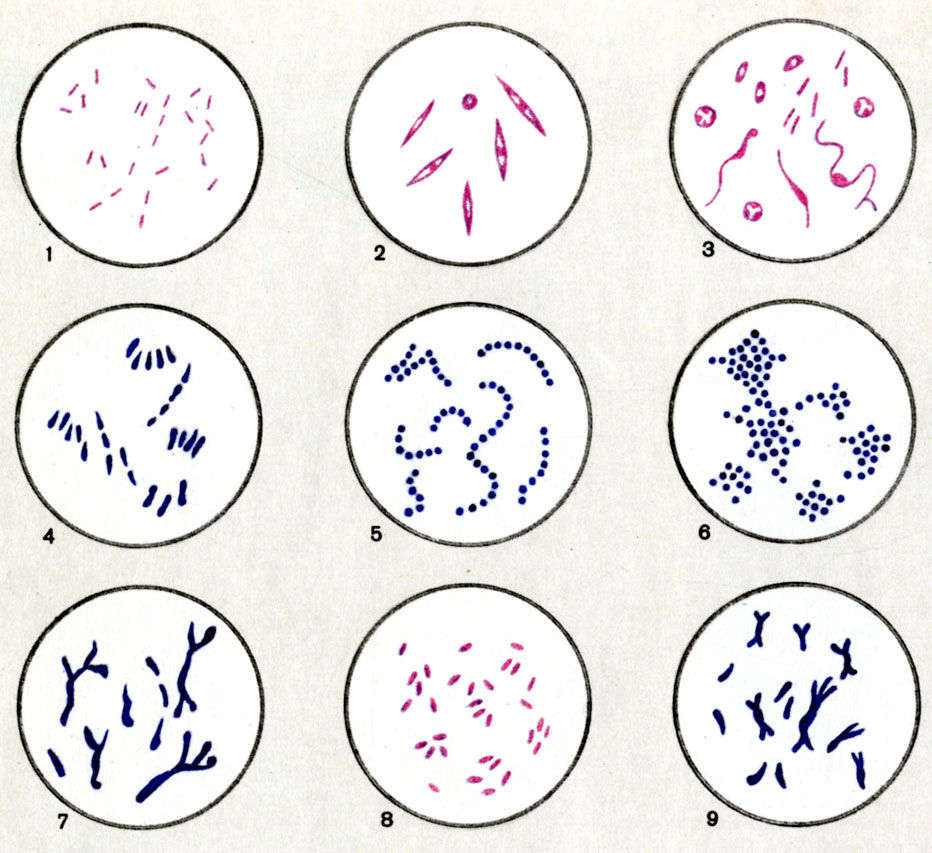
Морфология неспорогенных анаэробов (Прево, Тюрпен, Кайзер). Рис. 1. Bacteroides. Рис. 2. Fusobacterium. Рис. 3. Sphaerophorus. Рис. 4. Corynebacterium. Рис. 5. Peptostreptococcus. Рис. 6. Peptococcus. Рис. 7. Actinomyses. Рис. 8. Dilalister Рис. 9. Lactobacillus bifermentans
В 1861 г. Пастер впервые доказал, что нек-рые дрожжи и бактерии могут существовать и размножаться только при отсутствии воздуха (см. Анаэробиоз). Они были названы Пастером анаэробами.
Бактерии удовлетворяют свои потребности в энергии за счет сопряженных окислительно-восстановительных реакций, в ходе к-рых водород переносится от донатора к акцептору. У А. акцепторами водорода являются промежуточные продукты расщепления углеводов и белков, а у аэробов акцептором водорода может быть кислород. Эти реакции протекают ступенчато от системы более высокого окислительно-восстановительного потенциала к системе с более низким потенциалом. Перенос водорода от донатора к акцептору осуществляется четырьмя дегидрогеназами, причем в трех случаях в этом переносе в качестве акцепторов водорода участвуют пиридиннуклеотиды и только в случае сукцинатдегидрогеназы водород непосредственно переносится на флавопротеид (ФАД). По отношению к кислороду А. разделяются на две группы: факультативные и облигатные.
Факультативные анаэробы размножаются как в аэробных, так и в анаэробных условиях, в последнем случае в качестве акцептора водорода используют легко восстанавливающиеся элементы и соединения. Напр., многие анаэробные бактерии растут без кислорода, используя в качестве конечного акцептора электронов нитраты (нитратное дыхание). В этом случае, как показано Назоном (A. Nason, 1962) у Е. coli, перенос электронов осуществляется нитратредуктазой. У Cl. aceticum в качестве акцептора электрона служит углекислота. В анаэробных условиях метаболизм факультативных А. протекает по типу брожения, и субстрат полностью не окисляется. В присутствии кислорода происходит полное окисление субстрата, в результате чего высвобождается большое количество энергии и рост бактерий становится более интенсивным.
Облигатные анаэробы не способны усваивать кислород, в присутствии его они погибают. У одних облигатных А., включая представителей рода Clostridium, отсутствуют цитохромы, цитохромоксидазы, т. е. не происходит перенос водорода к кислороду. У других же А. цитохромоксидазы, к-рые переносят на молекулу кислорода два водородных иона, имеются,но это приводит к образованию перекиси водорода, токсичной для бактерий.Расщепление последней в клетке с выделением воды осуществляется двумя ферментами — каталазой и пероксидазой. В их отсутствие концентрация перекиси водорода достигает уровня, убивающего клетку. Поэтому облигатные А. можно культивировать только в среде, лишенной кислорода.
Нек-рые А., не имеющие ката лазы и цитохромов, могут в присутствии донаторов электронов — флавопротеинов — катализировать восстановление перекиси водорода до воды, в связи с этим перекись водорода у них не накапливается и не наступает гибель в присутствии кислорода.
А. играют большую роль в круговороте веществ, участвуя в разложении органических остатков растительного и животного происхождения без доступа воздуха или при затрудненном притоке его. При участии А. происходят процессы гниения в глубоких слоях почвы, в болотах, в иле, в навозных кучах. А. присутствуют в кишечнике человека и животных, участвуя в разложении растительной клетчатки. В среде, хорошо доступной для воздуха, А. принимают участие в разложении paзличных веществ совместно с аэробами, т. к. последние поглощают кислород.
Рис. 1-28. Анаэробы рода Clostridium. Рис. 1. Колонии столбнячной палочки (на агаре с мартеновским бульоном) с неровной, выпуклой зернистой поверхностьюс отростками (× 10)

Рис. 1-28. Анаэробы рода Clostridium. Рис. 2. Морфология барабанных палочек Cl. tetani (окраска по Рграму; × 1900)
Рис. 1-28. Анаэробы рода Clostridium. Рис. 3. Зона гемолиза вокруг колонии Cl. botulmum А на агаре с кровью (× 8)

Рис. 1-28. Анаэробы рода Clostridium. Рис. 4. Клетки Cl. Botulinum А (окраска по Граму; × 1900)
Рис. 1-28. Анаэробы рода Clostridium. Рис. 5. Морфология колонии Cl. brfulmff В на печеночном агаре (× 10)
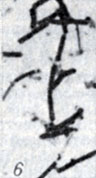
Рис. 1-28. Анаэробы рода Clostridium. Рис. 6. Морфология палочек Cl. botulinum В (окраска по Грму; × 1900)

Рис. 1-28. Анаэробы рода Clostridium. Рис. 7. Колонии Cl. botulinum C на агаре с бульоном Хоттингера (× 5)

Рис. 1-28. Анаэробы рода Clostridium. Рис. 8 Морфология пачочек и ракеток Cl. botulinum C (окраска фуксином; × 1900)
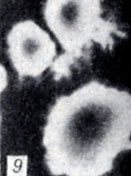
Рис. 1-28. Анаэробы рода Clostridium. Рис. 9. Колонии Gl. botulinum E на агаре с бульоном Хоттингера (× 5)

Рис. 1-28. Анаэробы рода Clostridium. Рис. 10. Морфология ракеток Cl. botulinum E (окраска по Граму; × 1900)
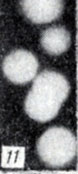
Рис. 1-28. Анаэробы рода Clostridium. Рис. 11. Колонии Gl. botulinum F на агаре с бульоном Хоттингера, выделенных из помета водоплавающих птиц с птичьего базара Баренцева моря (× 1900)
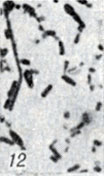
Рис. 1-28. Анаэробы рода Clostridium. Рис. 12 Морфология палочек и ракеток Cl. botulinum F, выделенных из помета водоплавающих птиц (окраска по Гюаму; × 1900)

Рис. 1-28. Анаэробы рода Clostridium. Рис. 13. Морфология клеток Cl. perfringens А из колоний на кровяном агаре (окраска фуксином; × 1900)
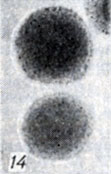
Рис. 1-28. Анаэробы рода Clostridium. Рис 14. Колонии Cl. perfringens B на агаре с бульоном Хоттингера (× 10)
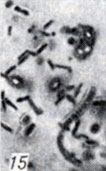
Рис. 1-28. Анаэробы рода Clostridium. Рис. 15. Морфология спор и палочек Cl. perfringens В в песке (окраска фуксином; × 1900)
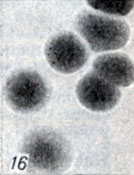
Рис. 1-28. Анаэробы рода Clostridium. Рис. 16. Колонии Cl. perfringens D на агаре с бульоном Хоттингера (× 10)

Рис. 1-28. Анаэробы рода Clostridium. Рис. 17. Морфология палочек Cl. perfrinlgens D (окраска по Граму; × 1900)
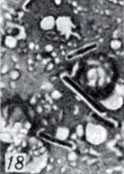
Рис. 1-28. Анаэробы рода Clostridium. Рис. 18. Капсула у Gl. perfringens А окюущает кчетку микпоба в виде светлой полоски. Мазок из печени морской свинки, погибшей от анаэробной инфекции (окраска фуксином; × 1000)
Рис. 1-28. Анаэробы рода Clostridium. Рис. 19. Колонии Cl. perfringens A на кровяном агаре окружены зоной гемолиза (натуральная величина)

Рис. 1-28. Анаэробы рода Clostridium. Рис. 20. Колонии Cl. oedematiens A на печеночном агаре (× 32)

Рис. 1-28. Анаэробы рода Clostridium. Рис. 21. морфология палочек Cl. oedematiens А со жгутиками (окраска по М. А. Морозову; × 1000)
Рис. 1-28. Анаэробы рода Clostridium. Рис. 22. Колонии Cl. septicum на печеночном агаре через сутки после посева (× 32)

Рис. 1-28. Анаэробы рода Clostridium. Рис. 23. Морфология клеток Cl. septicum (окраска фуксином; × 1900)

Рис. 1-28. Анаэробы рода Clostridium. Рис. 24. Cl. septicum в виде длинных нитей в печени морской свинки, погибшей от анаэробной инфекции (окраска по Грамму; × 1800)
Рис. 1-28. Анаэробы рода Clostridium. Рис. 25. Колонии Cl. sordelli на агаре (× 10)
Рис. 1-28. Анаэробы рода Clostridium. Рис. 26. Морфология клеток Cl. sordellii (окраска фуксином; × 1900)
Рис. 1-28. Анаэробы рода Clostridium. Рис. 27. Колония Cl. histolyticum на кровяном агаре (× 64)

Рис. 1-28. Анаэробы рода Clostridium. Рис. 28. Морфология Cl. histolyticum окоаска фуксином; × 1900)
По Берги (D. H. Bergey, 1957), факультативными или облигатными А. являются 93 вида спорогенных бактерий рода Clostridium, из к-рых более 10 видов патогенны для человека и животных. Эти бактерии (рис. 1—28) вызывают у людей следующие заболевания: Cl. tetani — столбняк (см.), Gl. botulinum — ботулизм (см.), Gl. perfringens, Gl. oedematiens, Cl. septicum, Cl. histolyticum, Cl. sordellii, Cl. fallax, Cl. chauvoei, Cl. sporogenes — анаэробную инфекцию (см.).
К облигатным или факультативным А. принадлежат и многие неспорогенные бактерии, грибки, а также трепонемы.
По классификации Берги (1957), к А. относятся девять родов: Bacteroides, Fusobacterium, Sphaerophorus, Corynebacterium, Peptostreptococcus, Peptococcus, Actinomyces, Dialister, Lactobacillus bilidus, всего 92 вида микроорганизмов.
По классификации Прево (A R Prevot, 1955, 1967), неспорогенные А. (цветн. табл., смотри выше, рис. 1—9) разделяются на 21 род, содержащий ИЗ видов, из к-рых более 33 видов патогенны для человека и животных. Особенно много патогенных видов анаэробов для человека среди родов Bacteroides, Sphaerophorus, Corynebacterium, Streptococcus, Staphylococcus, Actinomyces и др. Неспорогенные А. вызывают у человека гнойный плеврит, абсцесс легкого, послеродовой сепсис, послеабортный сепсис, инфицирование огнестрельных ран, перитонит, септицемию, абсцесс почек, мозга, печени, хрон. колит и другие заболевания,
Многие виды неспорогенных А. являются облигатными А. Выделение их из организма человека и животных весьма затруднено; при первых же пересевах на специальные среды они погибают от кислорода. Поэтому Хангейт (R. Е. Hungate, 1950) предложил облигатных А. пересевать и культивировать на восстановленных средах в атмосфере азота.
Для выращивания А. предлагалось много различных способов, обеспечивающих удаление кислорода из среды культивирования. Широко применяют аппараты, к-рые после удаления воздуха заполняют азотом (см. Анаэростат). Для поглощения кислорода при выращивании А. предложены хим. средства (напр., смесь пирогалловой к-ты с раствором NaOH или КОН).
Простой способ ограничения доступа воздуха в среду для А. — использование высокого слоя среды. Жидкую среду наливают в высокие сосуды (бутыли, высокие пробирки, флаконы), покрывают до стерилизации или после нее жидким вазелином, стерилизуют, перед посевом кипятят 20—30 мин. для удаления воздуха, быстро охлаждают и вносят посевной материал на дно сосуда. Для создания условий роста А. в среду вносят восстанавливающие вещества: глюкозу, аскорбиновую к-ту, муравьинокислый натрий, кусочки свежих тканей, вещества, содержащие SH-группу (тиогликолевую к-ту, цистеин, глутатион и др.), кусочки паренхиматозных органов животных, растительные ткани, культуры убитых дрожжей. Эти вещества должны вводиться в количествах, не угнетающих рост А.
Для выделения чистой культуры А. применяют метод Виньяля—Вейона. Пробирки (пипетки) длиной 20—30 см, диам. 6—7 мм заполняют на 15—20 см прозрачным 1—1,5% агаром. Перед посевом агар расплавляют, остужают до t° 40—50°, необломанную пастеровскую пипетку погружают в посевной материал, а затем вносят его поочередно в 5—7 пробирок (пипеток), агар быстро охлаждают. В глубине агара вырастают отдельные колонии А. в виде хлопьев, к-рые извлекают тонкой пипеткой либо разрезают пробирку (пипетку) и также достают отдельные колонии пастеровской пипеткой. Можно немного подогреть пробирку с агаром и перенести столбик в чашку Петри. Отдельные колонии пересевают на жидкие среды. Чистую культуру А. получают по методу Л. Г. Перетца, для чего запаянную пастеровскую пипетку предварительно погружают в посевной материал, а затем вносят поочередно в три пробирки, содержащие остуженный агар. К агару добавляют 2—4 капли 10% гипосульфита натрия на 10% растворе углекислой соды или 0,1 мл 8% раствора аскорбиновой к-ты на 10% растворе углекислой соды. Каждую пробирку выливают в стерильную чашку Петри, на дне к-рой лежит стекло размером 6×6 см на спичках или кусочках стекла. Агар с посеянным А. затекает под стекло, где вырастают отдельные колонии. Эти колонии можно пересеять на жидкую среду.
См. также Бактерии.
Виблиогр.: Матвеев К. И. Ботулизм М., 1959, библиогр.; он же, Эпидемиология и профилактика столбняка, М., 1960; Матвеев К. И. и Волгин Ю. Б. Анаэробная инфекция, Многотомн. руководство по микр., клин, и эпид. инфекц бол., под ред. Н. Н. Жукова-Вережникова, т. 7, с. 565, М., 1966; Мельников В. Н. и Мельников Н. И. Анаэробные инфекции, М., 1973, библиогр.; Bergey's manual of determinative bacteriology, ed. by R. S. Breed а. о Baltimore, 1957; Prévot A. R. Biologic des maladies dues aux anaérobies, P., 1955; Prévot A. R., Тurpin A. et Kaiser P. Les bactéries anaérobies, P., 1967.
Источники:
- Большая медицинская энциклопедия. Том 1/Главный редактор академик Б. В. Петровский; издательство «Советская энциклопедия»; Москва, 1974.- 576 с.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://sohmet.ru/ 'Sohmet.ru: Библиотека по медицине'