АМИГДАЛОИДНАЯ ОБЛАСТЬ
Расстановка ударений: АМИГДАЛО`ИДНАЯ О`БЛАСТЬ
АМИГДАЛОИДНАЯ ОБЛАСТЬ [лат. amygdaloideus — миндалевидный; син.: миндалевидное (амигдалоидное) тело (corpus amygdaloideum), миндалевидное (амигдалоидное) ядро (nucleus amygdalae), миндалина, миндалевидный (амигдалоидный, амигдалярный) ядерный комплекс, амигдала] — сложный комплекс ядер головного мозга, относящийся к базальным ядрам (архистриатум), участвующий в осуществлении корригирующего влияния на деятельность стволовых образований мозга.
Филогенетически А. о. — очень древнее образование мозга. Зачатки его можно проследить еще у круглоротых, но как самостоятельное ядро оно впервые появляется у амфибий и в дальнейшем сохраняется у всех позвоночных. В процессе онтогенеза А. о. формируется не только из ганглиозных (подкорковых), но и корковых формаций мозга, что отражается на общем характере ее строения.
Основная часть А. о., собственно миндалина по Брокхаусу (Н. Brockhaus, 1939), развивается из задневентральной части ганглионарного бугорка конечного мозга (так наз. примитивный стриатум) и древней коры (периамигдалоидной), покрывающей его и нередко простирающейся в глубь массы ядер. Кроме того, в образовании наружной и передней частей А. о. принимают участие определенные группы клеток ограды; дорсальная поверхность А. о. формируется при участии основного (матриксного) материала промежуточного мозга и ложа нижнего рога бокового желудочка.
Морфология. У млекопитающих А. о. мозга представляет собой скопление серого вещества, залегающего в глубине височной доли. Среди базалъных ядер (см.) А. о. занимает самое вентральное положение, являясь как бы продолжением чечевичного ядра (рис. 1). Под влиянием развития и увеличения объема новой коры у высших млекопитающих, и прежде всего у человека, ядра А. о. смещались, и в результате у человека А. о. располагается в передне-верхней части парагиппокампальной извилины, преимущественно в области крючка. Здесь масса ядер А. о. очень тесно соприкасается с древней корой, к-рая поэтому в этой области и названа периамигдалоидной. Т. о., у человека с нижней и медиальной поверхности А. о. граничит с периамигдалоидной и энторинальной областью коры. Снаружи А. о. соприкасается с белым веществом височной доли, наверху — с нижним краем ограды и чечевичным ядром, сзади простирается до хвоста хвостатого ядра и нижнего рога бокового желудочка, а около вершины этого рога она близко подходит к гиппокампу (см. Архитектоника коры головного мозга, Головной мозг).
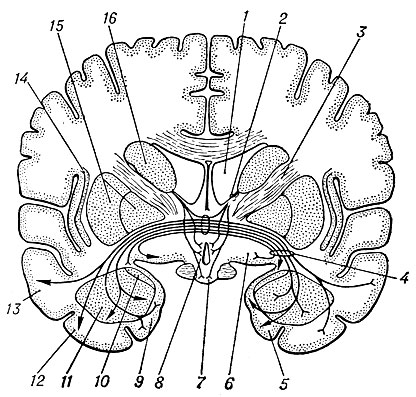
Рис. 1. Схема фронтального среза мозга человека на уровне конечного мозга; изображены все базальные ганглии (по Кросби, Хамфри, Лауэру): 1 — боковой желудочек; 2 — терминальная полоска; 3 — внутренняя капсула; 4 — передняя спайка; 5 — парагиппокампальная извилина; 6 — переднее продырявленное вещество; 7 — гипоталамус; 8 — преоптическая область; 9 — крючок; 10 — кортико-медиальный комплекс ядер амигдалоидной области; 11 — базо-латеральный комплекс ядер амигдалоидной области; 12 — нижняя височная извилина; 13 — средняя височная извилина; 14 — ограда (ciaustrum); 15 — чечевицеобразное ядро (бледный шар и скорлупа); 16 — хвостатое ядро
На основании цитологических особенностей А. о. можно разделить на несколько ядер. По данным сравнительной анатомии, у всех млекопитающих А. о. подразделяется на ядра. Однако до наст, времени классификация и номенклатура их окончательно не установлена. Существует несколько классификаций. Джонстон (1. Johnston, 1923) предложил выделить два комплекса ядер: кортико-медиальный и базо-латеральный (рис. 2). Кортико-медиальный комплекс составляют центральное, медиальное и корковое ядра, ядро латерального обонятельного тракта и кортико-амигдалоидная переходная зона. К базо-латеральному комплексу (филогенетически более молодому) относятся латеральное, базальное и дополнительное базальное ядра. При этом базальное ядро в свою очередь подразделяется на латеральную крупноклеточную и медиальную мелкоклеточную части. Впереди эта масса ядер образует так наз. переднюю А. о. Такая классификация остается до сих пор наиболее популярной в физиологической литературе.
Кросби и Хамфри (Е. Crosby, Т. Humphrey, 1941) в А. о. человека выделяют еще вставочное ядро и ядро терминальной полоски. Кроме того, латеральное ядро они подразделяют на пять частей: нежную, пограничную, крупно-, средне- и мелкоклеточную. Медиальную часть базаль-ного ядра они выделяют как отдельное вентро-медиальное ядро, а латеральную — как дорсо-латеральное, состоящее из четырех частей: крупно-, средне-, мелко- и микроклгеточной. Центральное ядро подразделяют на медиальную (крупноклеточную) и латеральную (микроклеточную) области.
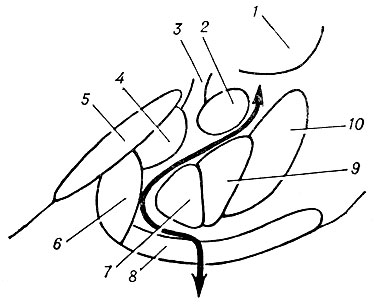
Рис. 2. Схематическое изображение ядер амигдалоидной области; жирная линия со стрелками показывает границу между базо-латеральным (справа) и кортико-медиальным (слева) комплексами ядер (по Коикегами): 1 — скорлупа; 2 — центральное ядро; 3 — терминальная полоска; 4 — медиальное ядро; 5 — зрительный тракт; 6 — корковое ядро; 7 — медиальная часть базального ядра; 8 — периамигдалоидная кора; 9 — латеральная часть базального ядра; 10 — латеральное ядро
В практике современной нейрохирургической клиники широко пользуются классификацией ядер А. о., предложенной Брокхаусом. Согласно этой классификации, корковое ядро А. о. называется поверхностным периамигдалоидным, медиальное — поверхностным надамигдалоидным, центральное — дорсальным глубоким надамигдалоидным, а латеральное — латеральным глубоким; латеральная часть базального ядра — промежуточным глубоким; медиальная часть базального ядра — вентральным глубоким и дополнительное базальное ядро — медиальным глубоким ядрами собственно амигдалы. Именно эта классификация ядер А. о. приводится в наиболее полном стереотаксическом атласе мозга человека, разработанном под руководством Шальтенбрандта и Бейли (G. Schaltenbrandt, P. Bailey, 1959).
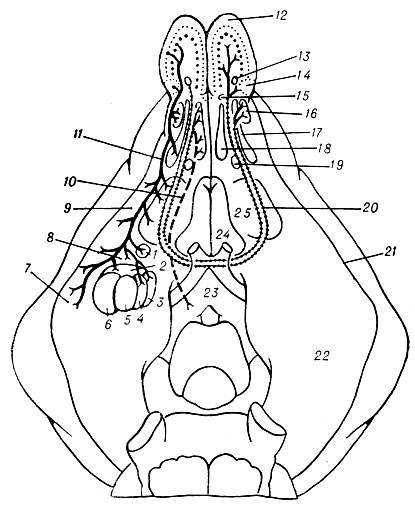
Рис. 3. Схема принципиальных связей обонятельной луковицы и переднего обонятельного ядра (по Лохману и Ламмерсу): 1 — ядро латерального обонятельного тракта А. о.; 2 — корковое ядро А. о.; 3 — медиальное ядро А. о.; 4 — центральное ядро А. о.; 5 — базальное ядро А. о.; 6 — латеральное ядро А. о.; 7 — пери-амигдалоидная кора; 8 — передняя амигдалоидная область; 9 — препириформная кора; 10 — медиальный пучок переднего мозга; 11 — латеральный обонятельный тракт; 12 — обонятельная луковица; 13 — ростральная часть переднего обонятельного ядра; 14 — дополнительная обонятельная луковица; 15 — дорсальная часть переднего обонятельного ядра; 16 — внешняя часть переднего обонятельного ядра; 17 — латеральная часть переднего обонятельного ядра; 18 — медиальная часть переднего обонятельного ядра; 19 — задняя часть переднего обонятельного ядра; 20 — обонятельная часть передней спайки; 21 — ринальная борозда; 22 — пириформная доля; 23 — гипоталамус; 24 — септальная область; 25 — обонятельный бугорок
Основные восходящие пути амигдалоидной области. В физиологии наиболее хорошо изучены и анатомически подтверждены амигдалопетальные волокна; самыми мощными из них являются волокна латерального обонятельного тракта, к-рые представляют собой аксоны митральных и пучковых клеток обонятельной луковицы. Амигдалопетальные волокна начинаются в обонятельной луковице, переднем обонятельном ядре, обонятельном бугорке и периамигдалоидной коре, идут к А. о. той же стороны (ипсилатерально) и оканчиваются в ее корковом и медиальном ядрах, а также в ядре латерального обонятельного тракта (рис. 3). По мнению Южлена (A. Hugelin), эти волокна оканчиваются и в медиальной части базального ядра. Гасто и Ламмерс (Н. Gastaut, H. J. Lammers) полагают, что волокна латерального обонятельного тракта направляются к А. о. противоположного полушария (контралатерально) и оканчиваются в центральном и базо-медиальном ядрах, а также в ядрах терминальных полосок А. о. обоих полушарий.
Другие амигдалопетальные проекции (зрительные, слуховые, вкусовые, соместетические и висцеростети-ческие пути), хотя и не подтверждены анатомически, выявлены с помощью электрофизиологических и нейроно-графических методов.
По-видимому, эти сенсорные амигдалопетальные связи осуществляются через таламические ядра, они были подтверждены анатомически и электрофизиологически. Что касается неспецифических ядер таламуса, то их проекции (пути) к А. о. окончательно не установлены, хотя нек-рые анатомические и электрофизиологические данные свидетельствуют о существовании таких проекций. Основная роль при этом отводится центральному медиальному ядру таламуса.
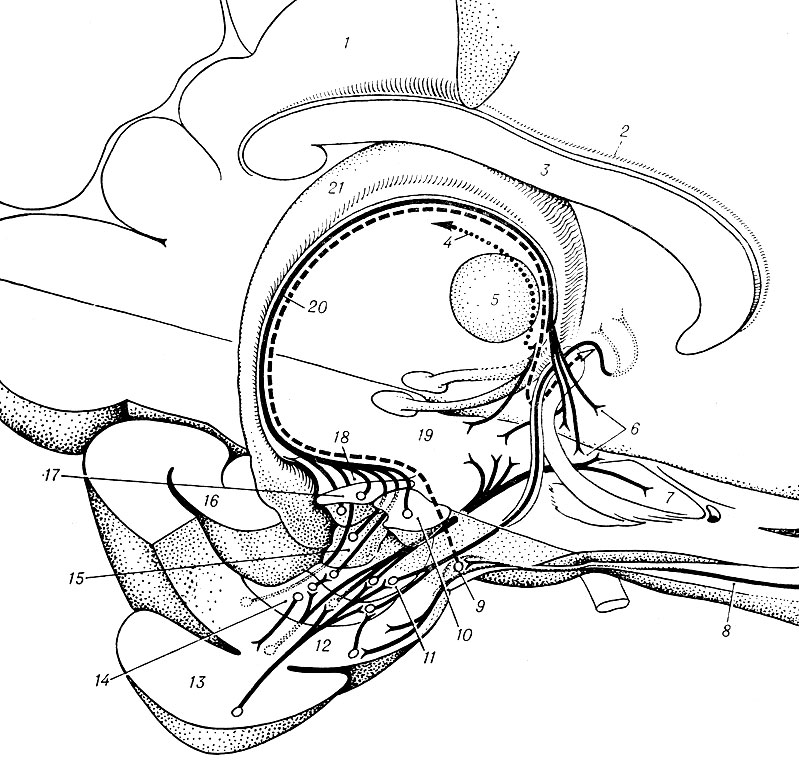
Рис. 4. Схематическое изображение структур височной доли, в том числе и ядер амигдалоидной области и ее основных (дорсальной и передневентральной) проводниковых систем (по Гасто и Ламмерсу): 1 — поясная извилина; 2 — серое покрывало; 3 — мозолистое тело; 4 — мозговая полоска; 5 — передние ядра таламуса; 6 — септальная область; 7 — nucleus accumbens; 8 — латеральный обонятельный тракт; 9 — ядро латерального обонятельного тракта; 10 — центральное ядро; 11 — базальное ядро; 12 — пириформная доля; 13 — височная доля; 14 — латеральное ядро; 15 — корковое ядро; 16 — гиппокамп; 17 — медиальное ядро; 18 — передняя амигдалоидная область; 19 — гипоталамус; 20 — терминальная полоска; 21 — свод
Важная особенность нейронов А. о. заключается в том, что они воспринимают импульсы, приходящие от различных органов чувств (поликонвергентность).
Обнаружены уздечко-амигдалоидные пути, проходящие в составе мозговой и терминальной полосок. Полагают, что уздечка осуществляет связь между А. о. обоих полушарий. Из других подкорковых образований, посылающих свои волокна к А. о., необходимо указать на ядра полосатого тела, ретикулярную формацию ствола мозга и гипоталамус. Ананд (В. Anand, 1959) с сотр. обнаружил мозжечково-амигдалоидные связи.
А. о. воспринимает проекции и от формаций старой, древней и новой коры. Кроме волокон от уже упомянутого обонятельного бугорка, она получает много волокон от коры парагиппокампальной извилины, особенно ее передней части. По мнению большинства исследователей, эти волокна оканчиваются во многих ядрах А. о. Гиппокамп, по-видимому, тоже посылает волокна к А. о., хотя и меньше, чем пириформная кора. Анатомически установлены височно-амигдалоидные связи (рис. 4), приходящие к А. о. от новой коры. Уитлок и Наута (D. Whitlock, W. Nauta, 1956) обнаружили проекции нижней височной извилины к базо-латеральному комплексу ядер А. о., а Сегундо (J. P. Segundo, 1955) с сотр. показал, что эти ядра получают импульсы и от верхней височной извилины. Было обнаружено, что волокна полюса височной доли оканчиваются в латеральном и центральном ядрах А. о.
Кортикофугальные волокна от передней орбитальной коры до латерального ядра А. о. проследил Коикегами (Н. Koikegami, 1963). Прослежены также волокна от полюсов фронтальной и височной долей коры до базо-латерального комплекса А. о. С помощью метода физиологической нейронографии были установлены пути, идущие к А. о. от задней орбитальной и передней инсулярной областей, а также от полюса височной доли и от моторной коры. Относительно проекций в А. о. коры поясной извилины единого мнения не существует.
Основные нисходящие пути амигдалоидной области. Среди множества отходящих от А. о. путей можно выделить две главные и наиболее изученные мощные системы: дорсальную (терминальную полоску) и передне-вентральную (рис. 4 и 5).
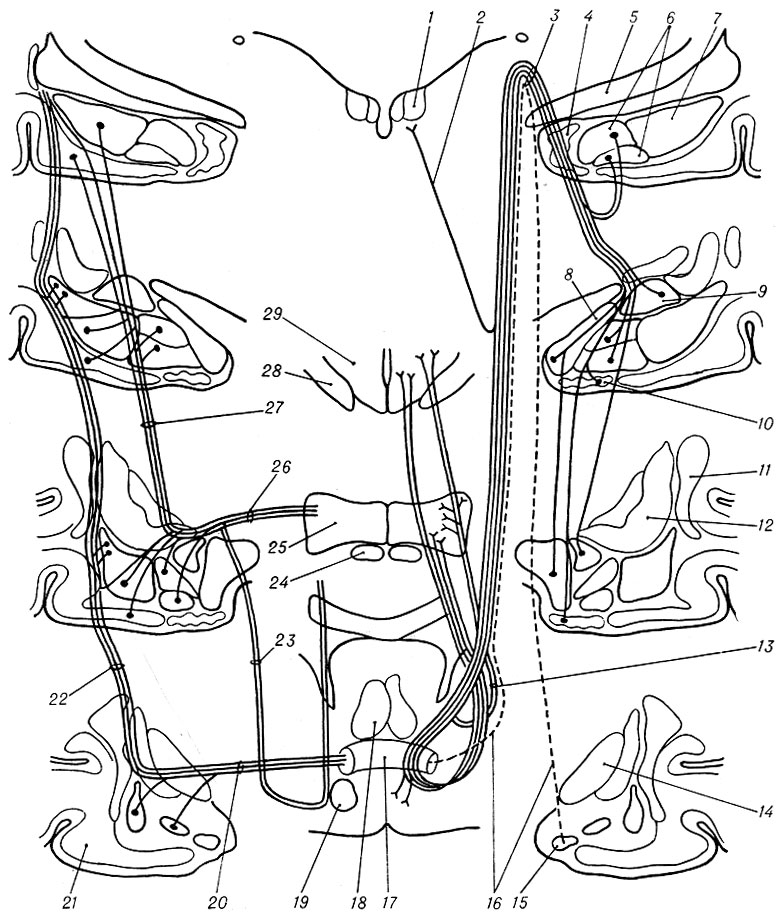
Рис. 5. Схема основных волоконных связей амигдалоидной области; справа изображена терминальная полоска и различные структуры мозга, связанные с ее отдельными компонентами, слева — внешняя капсула, передняя подкова передней спайки, продольный ассоциативный пучок и структуры мозга, связанные с этими проводящими путями (по Фоксу): 1 — уздечковый ганглий; 2 — мозговая полоска (часть терминальной полоски); 3 — терхминальная полоска со всеми ее компонентами; 4 — гиппокамп; 5 — зрительный тракт; 6 — базальное ядро; 7 — латеральное ядро; 8 — медиальное ядро; 9 — центральное ядро; 10 — корковое ядро; 11 — ограда; 12 — скорлупа; 13 — преоптический компонент терминальной полоски; 14 — бледный шар; 15 — ядро латерального обонятельного тракта амигдалоидной области; 16 — комиссуральный компонент терминальной полоски; 17 — передняя спайка; 18 — колонны свода; 19 — медиальный пучок переднего мозга; 20 — передняя подкова передней комиссуры; 21 — пириформнаякора; 22 — внешняя капсула; 23 — передняя часть продольного ассоциативного пучка; 24 — зрительный нерв; 25 — преоптиче-ская область; 26 — задняя часть продольного ассоциативного пучка; 27 — продольный ассоциативный пучок в целом; 28 — зрительный тракт; 29 — гипоталамус
Терминальная полоска представляет собой четко очерченный компактный пучок волокон, проходящий над внутренней капсулой и разделяющийся в районе передней спайки на четыре отдельных компонента: комиссуральный, супра-, инфра- и посткомиссуральный. Волокна комиссурального компонента идут от ядра латерального обонятельного тракта и, возможно, от медиального и центрального ядер А. о. (на рис. 4 этот компонент полоски обозначен пунктирной линией). Около передней спайки его волокна отделяются от терминальной полоски и в составе задней дуги спайки переходят на противоположное полушарие, где смешиваются с волокнами терминальной полоски и, по мнению нек-рых авторов, оканчиваются в ядре латерального обонятельного тракта А. о. другой стороны или в преоптических областях обоих полушарий. Наиболее важным в морфологическом и функциональном отношении представляется супракомиссураль-ный компонент терминальной полоски. Его волокна возникают в задне-базальном и кортико-медиальном комплексах ядер А. о., а также в ее центральном ядре и оканчиваются в ядре-ложе терминальной полоски (к-рое достигает своего наибольшего развития в области передней спайки), прекомиссуральной части септальной области и ее п. accumbens и значительная часть — в преоптической области (рис. 5). Пост- и инфракомиссуральные компоненты волокон терминальной полоски часто рассматриваются вместе и объединяются под общим названием «преоптикогипоталамический компонент», поскольку они оканчиваются гл. обр. в гипоталамусе и преоптической области. Начало этим волокнам, как и волокнам супракомиссурального компонента, дают одни и те же ядра А. о. Значительная часть этих волокон оканчивается в вентро-медиальном и перивентрикулярных ядрах гипоталамуса (рис. 5). От посткомиссурального компонента терминальной полоски в области переднего отдела таламуса отделяется уздечковый пучок, или мозговая полоска (на рис. 4 она представлена в виде точечной линии). Волокна этого пучка связывают центральное и корковое ядра А. о., периамигдалоидную кору, самый каудальный отдел А. о., пограничный с гиппокампом, и уздечковый ганглий.
Передне-вентральная нисходящая система волокон А. о. изучена гораздо меньше. Это полиморфная волоконная сеть, включающая большое число диффузных мультисинаптических путей, конечные пункты к-рых часто совпадают с конечными пунктами терминальной полоски (рис. 5).
Различные авторы называют эту амигдалофугальную волоконную организацию по-разному: продольный ассоциативный пучок, амигдало-пириформный ассоциативный пучок, латеральный кортико-гипоталамический пучок, пучок вентральной обонятельной проекции и т. д. Амигда-лофугальные волокна начинаются в основном в пириформной области коры и базо-латеральном комплексе ядер А. о., направляются вперед, проходя под височной «подковой» передней спайки и оставляя на пути коллатерали в септальной, передней амигдалоидной областях, пириформной области коры, обонятельном бугорке, переднемозговом пучке, переднем обонятельном ядре, безымянной субстанции, и оканчиваются в латеральных частях преоптической области и гипоталамуса. Кроме этих сложных нисходящих путей, нек-рые исследователи выявили связи А. о. с энтопедункулярным ядром и таламусом (подушкой, дорсо-латеральным и задне-латеральным ядрами). У птиц и нек-рых рептилий установлены амигдало-мезэнцефалические проекции.
Из областей коры, воспринимающих импульсы, исходящие из А. о., прежде всего нужно назвать пириформную, к-рая является одновременно и переключательной станцией на их пути к гиппокампу, с к-рым прямые анатомические связи не установлены. Однако эти связи были выявлены с помощью нейронной стрихнинографии. Электрофизиологические приемы исследования позволили проследить проекции от А. о. к коре височной области, островку и нек-рым участкам коры лобной области.
Между собой А. о. обоих полушарий связаны волокнами передней спайки.
Физиология амигдалоидной области. Будучи одной из главных структур лимбической системы (см.), А. о. участвует в осуществлении корригирующего влияния на деятельность стволовых образований мозга, где локализуются центры жизненно важных интегративных реакций организма.
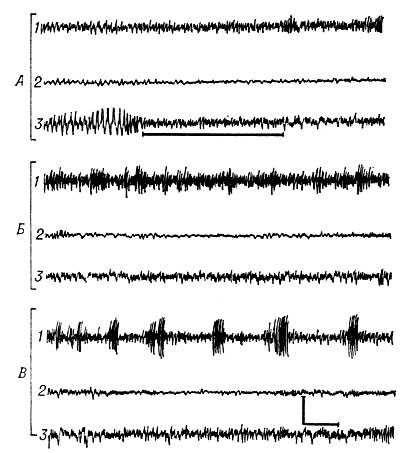
Рис. 6. Изменение фоновой электрической активности в различных структурах мозга кошки (по Ониани): 1 — базальное ядро амигдалоидной области; 2 — латеральное ядро гипоталамуса; 3 — височная (новая) кора; А — при простой ориентировочной реакции на тон; Б — при реакции настороженности в ответ на раздражение ретикулярной формации среднего мозга; В — во время реакции страха в ответ на раздражение латерального ядра гипоталамуса. Калибровка: 200 iмкв/i, 1 сек
Спонтанная биоэлектрическая активность А. о. характеризуется нерегулярными ритмическими колебаниями средней амплитуды (100—200 мкв) и частоты (4—8 кол/сек). Во время ориентировочной реакции организма, вызванной действием какого-либо внешнего стимула и раздражением ретикулярной формации (см.), на ее электрограмме почти синхронно с ритмом дыхания появляются вспышки быстрых ритмичных колебаний (40—55 кол/сек). Выраженность этих колебаний увеличивается под действием обонятельной стимуляции и раздражения ретикулярной формации мозга. Именно это отличает А. о. от новой коры, на электрограмме к-рой во время ориентировочной реакции животного (см. Ориентировочно-исследовательская реакция) регистрируется десинхронизация спонтанных биоэлектрических колебаний (рис. 6).
При раздражении А. о. электрическим током наблюдаются самые разнообразные интегративные реакции, свидетельствующие о том, что А. о. оказывает влияние на многие функции организма. Так, при раздражении А. о. даже слабым электрическим током у животного возникает типичная реакция пробуждения или ориентировочно-исследовательская реакция с элементами беспокойства (животное настораживается, прислушивается, зрачки расширяются, учащается ритм сердца, дыхание и т. д.), на электроэнцефалограмме — диффузная десинхронизация биоэлектрической активности новой коры. Сразу после удаления А. о. обоих полушарий у животных наблюдается сонливость и заметное снижение общей активности. Кошки, напр., подолгу сидят в одном положении, напоминающем позу сфинкса, температура тела значительно снижается.
А. о. активно влияет на формирование и других интегративных реакций организма, прежде всего на мотивационно-эмоциональную сферу (рис. 7). Участие А. о. в развитии мотивационных состояний наиболее отчетливо проявляется в пищедобывательном и половом поведении (см. Мотивации). Так, при раздражении А. о. электрическим током у животных наблюдаются целостные пище-добывательные реакции, как, напр., разыскивание и поедание пищи, и симптомы так наз. орального комплекса движений — слюноотделение, принюхивание, облизывание, жевание, глотание и т. д. Раздражение А. о. голодных животных, как правило, приводит к торможению пищевой активности: прекращается поиск, животное отказывается от пищи, значительно ухудшается выработка пищевых условных рефлексов. Двустороннее удаление А. о. приводит к уменьшению или увеличению потребления пищи. Иногда у одного и того же животного сначала развивается снижение пищевой активности, а затем ее усиление, причем последнее оказывается более стойким и длительным. Чаще в послеоперационном периоде у животных наблюдается тенденция к повышенному поглощению пищи и, соответственно, к более активному ее поиску.
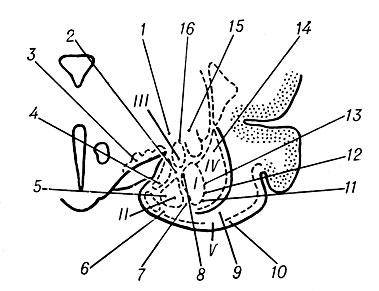
Рис. 7. Схема основных групп ядер амигдалоидной области, при раздражении которых возникает комплекс интегративных реакций (по Мак-Лейну и Дельгадо): 1 — глотание, мигание, поворот головы в сторону, противоположную стороне раздражения, сгибание задней конечности на стороне, противоположной стороне раздражения; 2 — жевание, рвотные движения, закрывание глаза на стороне раздражения, ускорение дыхания; 3 — ворчание, облизывание, жевание, поворот головы назад в сторону, противоположную раздражению, расширение зрачков, замедление ритма сердца, пилоэрекция, животное скребет контрлатеральной лапой; 4 — шипение, рычание, расширение зрачков, тяжелое дыхание; 5 — глотание, сокращение лицевых мышц на стороне раздражения, подавление дрожания и мурлыканья, учащение дыхания; 6 — фырканье, облизывание рта, закрывание ипсилатерального глаза, поворот головы в сторону, противоположную стороне раздражения, учащение дыхания; 7 — облизывание, жевание, закрывание глаза и сокращение лицевых мышц на стороне раздражения, поднимание головы; 8 — облизывание, мигание, закрывание ипсилатералъного глаза, прекращение мурлыканья; расширение зрачков, замедление или ускорение дыхания; 9 — жевание, облизывание, рычание, расширение зрачков, замедление дыхания; 10 — подергивание мышц лица, прижатие уха на стороне раздражения, мочеиспускание, животное кусается, скребет лапой и поворачивается в сторону, противоположную стороне раздражения, саливация, тяжелое дыхание; 11 — облизывание, замедление дыхания, животное огрызается в сторону, соответствующую стороне раздражения; 12 — жевание, ускорение дыхания, снижение частоты сердечных сокращений; 13 — подергивание ипсилатеральных мышц лица, тяжелое дыхание; 14 — пробуждение, настораживание ушей, расширение зрачков, подавление дыхания; 15 — облизывание, сокращение ипсилатеральных мышц лица; 16 — мяуканье, глотание, прижатие уха на стороне, противоположной стороне раздражения; разгибание ипсилатеральной передней лапы; поворот тела в сторону раздражения, замедление дыхания; I и II — латеральные и медиальные части базальных ядер миндалины; III — центральное ядро; IV — боковое ядро; V — грушевидная извилина
При осуществлении пищедобывательного поведения в А. о. кошек регистрируется характерная веретенообразная электрическая активность с частотой 35—40 колебаний в 1 секунду.
Пищевая активность животных тесно связана с функцией ядер базо-латерального комплекса А. о., хотя определенное участие принимает и кортико-медиальный комплекс. По-видимому, между этими комплексами существуют реципрокные взаимоотношения (см. Реципрокность). Напр., при двустороннем удалении базального ядра потребление и усвоение пищи настолько возрастают, что вес животных может увеличиваться на 20%.
Повреждение коркового ядра зачастую приводит к ослаблению пищевой активности. На основании сравнительных морфологических и функциональных данных многие исследователи предполагают, что существует система структур мозга, объединяющая А. о., септальную область и хвостатое ядро, к-рая параллельно с гипоталамусом осуществляет регуляцию пищевой активности.
А. о. оказывает значительное влияние и на половое поведение. Результаты многочисленных исследований, проведенных на разных видах животных, свидетельствуют о том, что в течение нескольких недель после двустороннего удаления А. о. животные бывают гиперсексуальными. Самцы пытаются копулировать с самцами, с животными других видов, с самками, не находящимися в состоянии течки, и даже с неодушевленными предметами. Гиперсексуальность животных в данных случаях прекращалась после кастрации и после разрушения вентро-медиального ядра гипоталамуса или ядер перегородки.
Иногда двусторонняя экстирпация А. о. приводит к ослаблению половой функции. У самок обезьян, как правило, рождается неполноценное, нежизнеспособное потомство, у самцов — понижается сперматогенез. Гиперсексуальность может наблюдаться и при раздражении А. о. электрическим током.
Описаны случаи, когда при атрофических нарушениях в А. о. мозга и при оперативных вмешательствах в этой области у человека возникали нек-рые патологические реакции, сходные с вышеописанными у животных; наблюдались расстройства, связанные с пищевым поведением, гиперсексуальность, приводящая больного, напр., к эксгибиционизму, мастурбации.

Рис. 8. Кошка, у которой удалены мигательные перепонки и левый шейный симпатический ганглий. В результате раздражения амигдалоидной области значительно расширяется правый зрачок и сужается левый (по Коикегами)
Наиболее выражено влияние А. о. на формирование у животных эмоциональных реакций, тесно связанных с мотивационными состояниями. Напр., у кошек, лишенных А. о. обоих полушарий, практически исчезает эмоциональный компонент пищедобывательной деятельности, в то время как собственно пищевая активность страдает гораздо меньше: способность поглощать поставленную рядом пищу полностью сохраняется.

Рис. 9. Замедление роста тела животных после двустороннего удаления всего комплекса ядер амигдалоидной области (по Коикегами): вверху — подопытная, внизу — контрольная крыса
Двустороннее удаление А. о. приводит к обеднению эмоциональной сферы животных. Дикие агрессивные животные перестают реагировать на человека и на животных других видов, даже на тех, на к-рых они обычно нападают. У них снижается реактивность на вредоносные и болевые раздражения (на ноцицептивные раздражения) и на сигналы об опасности.
Удаление А. о. у животных, к-рые в результате разрушения гипоталамуса и септальной области стали гиперэмоциональными, приводит к значительному ослаблению эмоциональности.
У животных с разрушенными базо-латеральными ядрами А. о. заметно нарушается выработка оборонительных условных рефлексов, тормозятся, а иногда и полностью исчезают все ранее выработанные условные рефлексы.
Для А. о. в целом характерен двузначный (амбивалентный) тип эмоционального функционирования.
Так, при раздражении А. о. электрическим током у животных наблюдаются эмоциональные реакции, в основном отрицательного знака — испуг, страх, агрессия, сопровождающиеся выраженными моторными, вегетативными и голосовыми компонентами. Описаны клинические наблюдения, когда стимуляция А. о. у человека, применяемая в терапевтических целях, иногда вызывала чувство страха, тоски и испуга. Вместе с тем раздражение А. о. может вызывать и положительные эмоции, к-рые более всего проявляются в виде реакции самораздражения (см.).
Иногда при раздражении одних и тех же точек А. о. у животных наблюдаются эмоциональные реакции обоих знаков. Решающую роль в этом случае играют, по-видимому, параметры электрического тока. Однако наиболее выраженные положительные реакции можно получить, стимулируя кортико-медиальный комплекс ядер, а наиболее отрицательные — базо-латеральный комплекс.
Двустороннее удаление А. о. приводит к нарушениям и во взаимоотношениях животных: оперированные особи начинают избегать общения с сородичами и другими животными стада, стараются спрятаться. Будучи разобщенными с сородичами, они, в отличие (от интактных животных, не пытаются вернуться в свое сообщество. Вожаки сообществ после операции переходили на подчиненное положение.
А. о. играет важную роль и в регуляции вегетативных и моторных реакций, представляющих собой внешние проявления аффективных состояний животных. Стимуляция электрическим током или повреждения А. о. изменяют дыхание, работу сердца и пищеварительных органов, уровень кровяного давления, сахара и гормонов в крови, ширину зрачка (рис. 8), кожно-гальваническую реакцию, пилоэрекцию (см.). Поскольку А. о. связана с симпатической и с парасимпатической нервной системой, изменения вегетативных функций неоднозначны. Однако при двустороннем удалении А. о. чаще можно наблюдать ослабление работы сердца и легких, повышение кровяного давления и уровня сахара в крови.
На основании многочисленных экспериментов было выяснено, что влияния А. о. на вегетативную нервную систему опосредуются, как правило, через гипоталамус (см.). Было установлено, что кортико-медиальный комплекс ядер А. о. оказывает большое влияние на симпатическую зону гипоталамуса, а базо-латеральный комплекс — на парасимпатическую зону. При экстирпации соответствующих ядер гипоталамуса эти влияния исчезают. Базо-латеральная группа ядер связана также с экстрапирамидной системой (см.) мозга, поэтому при раздражении А. о. электрическим током возникает целый ряд моторных расстройств.
А. о. оказывает влияние и на эндокринную систему организма. После двустороннего удаления А. о. деятельность эндокринной системы, как правило, резко понижается. Это особенно заметно, если операция проводится на молодых животных. У них замедляется рост тела (рис. 9), атрофируется гипофиз, надпочечники, поджелудочная и щитовидная железы. Электрической стимуляцией А. о. можно усилить секрецию 17-оксикор-тикостероидов, гонадотропного гормона, изменить моторику жел.-киш. тракта, нарушить течение беременности и т. д.
А. о. оказывает определенное влияние на высшую нервную деятельность, память и сенсорное восприятие. При повреждениях А. о. нарушаются ранее выработанные условные рефлексы (особенно дифференцировочные), затрудняется выработка новых рефлексов и становится очень сложной переделка сигнальных значений условных раздражителей [Н. М. Вавилова, 1967; Фонберг (Е. Fonberg), 1961]. Так, напр., обезьянам с поврежденным амигдалоидно-гиппокампальным комплексом для дифференцирования зрительных раздражителей требовалось в 4 раза больше, а для переделки сигнального значения раздражителей — в 6—7 раз больше попыток, чем интактным животным.
При двустороннем удалении А. о. понижаются пороги восприятия сенсорных сигналов, нарушается дифференцировка их интенсивности. Иногда у животных нарушается аккомодация глаз. Раздражение А. о. электрическим током усиливает конвергенцию различных сенсорных возбуждений на одном нейроне новой коры.
Существует мнение, что все указанные нарушения интегративных функций мозга, возникающие в результате какого-либо раздражения А. о., могут проявиться только в том случае, когда в А. о. в результате этого раздражения возникает биоэлектрическая гиперсинхронная судорожная активность.
Общеизвестно, что А. о. обладает низким порогом судорожных реакций.
Развивающаяся в А. о. в ответ на раздражение гиперсинхронная биоэлектрическая активность имеет тенденцию распространяться на соседние и отдаленные области, в результате чего состояние повышенной возбудимости охватывает многие стволовые структуры мозга и прежде всего гипоталамус, таламус, ретикулярную формацию, а также перегородку, полосатое тело и структуры архипалеокортекса (см. Архитектоника коры головного мозга, больших полушарий). Можно думать, что именно с этим связаны расстройства памяти, сенсорного восприятия, условнорефлекторной деятельности и другие мотивационно-эмоциопальные нарушения.
Клинические наблюдения показывают, что у 40% больных эпилепсией судорожный синдром содержал аффективный компонент (страх, тоску или сильную немотивированную депрессию); при этом эпилептический очаг у большинства наблюдаемых больных находился вблизи А. о.
Органические поражения А. о. мозга приводят к так наз. височной эпилепсии; эпилептические пароксизмы в этом случае проявляются симптомами психомоторного, вегетативного и эмоционального характера. У таких больных нарушаются многие основные мотивации, в результате чего происходит расстройство пищевых реакций (повышение или снижение аппетита), половой активности (гипер- или гипосексуальность), оборонительного поведения (приступы неудовольствия, немотивированного страха, злобности, ярости, а порой и агрессивности — см. Оборонительные реакции). Появление этих симптомов связано, как правило, с возникновением электросудорожной активности А. о. Однако электросудорожная активность А. о. не всегда связана с патологическими процессами. Многие исследователи рассматривают эту активность как нейрофизиологическую основу корригирующего влияния, к-рое А. о. в норме оказывает на структуры мозгового ствола.
Библиогр.: Крачун Г. П. Межцентральная организация функции миндалевидных ядер, в кн.: Физиол. и биохим. ядер мозга, под ред. А. М. Марица и др., с. 30, Кишинев, 1971, библиогр.; Мгалоблишвили М. М. Влияние повреждения миндалевидного комплекса на условно-рефлекторную активность и краткосрочную память животных, Сообщ. АН Груз. ССР, т. 62, № 3, с. 685, 1971, библиогр.; Филимонов И. Н. Строение миндалевидного ядра у человека и его изменения в процессе онто- и филогенеза, Вестн. АМН СССР, № 5, с. 37, 1958, библиогр.; Черкес В. А. К физиологии миндалевидных ядер, в кн.: Гагрские беседы АН Груз. ССР, под ред. И. С. Бериташвили, т. 5, с. 258, М., 1968, библиогр.; Шугалев Н. П. Изменения электрической веретенообразной активности в миндалевидном ядре мозга кошки при осуществлении пищедобывательного рефлекса, Журн. высш. нервн. деятельн., т. 23, в. 4, с. 738, 1973; Brockhaus H. Zur normalen und pathologischen Ana-tomie des Mandelkerngebietes, J. Psychol. Neurol. (Lpz.), Bd 49, S. 1, 1939, Bibliogr.; Druga R. Neocortical projections to the amygdala, J. Hirnforsch., Bd 11, S. 467, 1970; Egger M. D. a. Flynn J. P. Further studies on the effects of amygdaloid stimulation and ablation on hypothalamically elicited attack behavior in cats, Progr. Brain Res., v. 27, p, 165, 1967; Gastaut H. et Lammers H. J. Les grandes activites du rhinencephale, t. 1, P., 1961, bibliogr.; Glооr P. Amygdala, Handb. physiol., Sec. 1 — Neurophysiology, ed. by J. Field a. o., v. 2, p. 1395, Washington, 1960, bibliogr.; Jacobs B. L. а. Мс Gintу D. J. Participation of the amygdala in complex stimulus recognition and behavioral inhibition, Brain Res., v. 36, p. 431, 1972, bibliogr.; Jоnasоn K. R. a. Enlое L. J. Alterations in social behavior following septal and amygdaloid lesions in the rat, J. сотр. physiol. Psychol., v. 75, p. 286, 1971, bibliogr.; Magnus O. et Naquet R. Physiologic normale et patholo-gique de 1'amygdale, в кн.: Les grandes activites du rhinencephale, t. 2, p. 191, P., 1961, bibliogr.; Matheson G. K., Branch B. J. а. Taylor А. N. Effects of amygdaloid stimulation on pituitary-adrenal activity in conscious cats, Brain Res., v. 32, p. 151, 1971, bibliogr.; Niemer W. T. a. o. Thalamo-amygdaler relationships, ibid., v. 24, p. 191, 1970; Spevасk A. A. a. Pribrаm К. Н. Decisional analysis of the effects of limbic lesions on learning in monkeys, J. сотр. physiol Psychol., v. 82, p. 211, 1973; Vergnes M. et Кarli P. Effet des lesions amygdaliennes sur le comportement d'agression interspécifique provoque che2 le rat par des lésions hypothalamiques medianes, C. R. Soc. Biol. (Paris), t. 164, p. 2647, 1970, bibliogr.
Источники:
- Большая медицинская энциклопедия. Том 1/Главный редактор академик Б. В. Петровский; издательство «Советская энциклопедия»; Москва, 1974.- 576 с.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://sohmet.ru/ 'Sohmet.ru: Библиотека по медицине'