7.1.3.1. Стероидные гормоны, их химическая номенклатура, биосинтез и метаболизм
Наиболее важные для практики сексопатолога гормоны, осуществляющие непосредственную регуляцию половых функций, по химической природе относятся к группам: 1) белково-пептидных гормонов (рилизинг-гормоны гипоталамуса, стимулирующие выделение тропных гормонов передней доли гипофиза, а также большая часть этих гормонов - АКТГ, фолликулостимулирующий, лютеинизирующий, гормон роста и др.); 2) стероидных гормонов [вырабатываются в надпочечниках, яичках, яичниках и плаценте; это мужские половые гормоны (андрогены), женские половые гормоны (эстрогены и прогестины) и гормоны, выделяемые корой надпочечников (кортикостероиды)].
Все стероидные гормоны берут начало от углеводорода стерана, структурной основой которого является циклопентанпергидрофенантреновое ядро (рис. 15), образованное четырьмя соединенными между собой углеродными кольцами (А, В, С, D): три кольца состоят из 6, а четвертое - из 5 углеродных атомов.
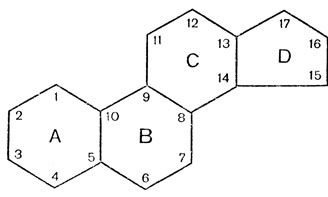
Рис. 15. Циклопентанпергидрофенантреновое ядро
Молекула циклопентанпергидрофенантрена содержит 17 атомов углерода; к стерану часто присоединяется различной длины боковая цепь, атомы углерода которой обозначаются цифрами, начиная с 18. Стероидные гормоны разделяются на три группы:
1. Эстран, образуемый путем присоединения к стерану метильной группы СН3, является материнским соединением для всех эстрогенных гормонов, в состав которых входят 18 атомов углерода,- группа С18 (рис. 16).
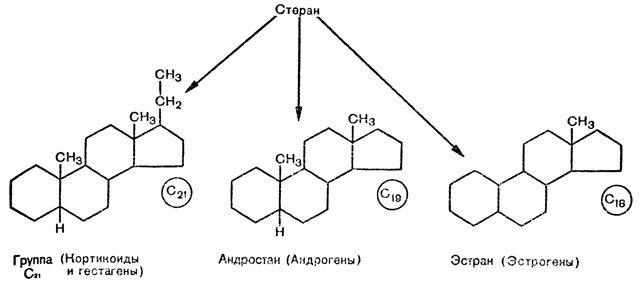
Рис. 16. Три основных (материнских) соединения: эстран (18 атомов углерода), из которого образуются эстрогены; андростан (19 атомов углерода), на основе которого образуются андрогены; группа C21 (21 атом углерода) - основа образования кортикоидов и гестагенов
2. Андростан, образуемый путем присоединения к стерану двух метильных групп, является материнским соединением для андрогенов (C19, рис. 16).
3. Группа с 21 атомом углерода (C21), к которой относятся гестагены (прогестероновые соединения), а также кортикоиды. Они содержат две метильные группы (CH3), а к 17-му атому углерода присоединена этильная группа CH2 - CH3 (см. рис. 16).
Обычно символ CH3, в частности у 10-го и 13-го атомов углерода, для упрощения не пишут, а обозначают лишь связь в виде черточки, направленной вверх.
Если в молекуле нет двойных связей, то название соединения имеет окончание "ан" (например, эстран, андростан и т. д.). Если в молекуле имеется двойная связь, то название приобретает окончание "ен" (при двух двойных связях - "диен", при трех - "триен"). Двойные связи также могут обозначаться Δ (при этом около буквы пишут цифры порядковый номер атома углерода, от которого начинается двойная связь).
Если водород, связанный с одним из атомов углерода, замещен кислородом с образованием гидроксильной (или алкогольной) группы, то название гормона оканчивается на "ол" (обозначает наличие группы ОН). При двух группах ОН окончание изменяется на "диол", при трех - на "триол". Если стероидные соединения содержат кетоновую группу С = О, то название оканчивается на "он" (при двух кетоновых группах - "дион", при трех - "трион").
Кроме названных обозначений пользуются и другими: "дезокси" - когда первичная молекула утратила атом кислорода, "дегидро" - когда молекула утратила два атома водорода.
Наконец, в номенклатуре стероидных гормонов встречаются приставки "гидрокси" или "окси", которые обозначают, что атом водорода в молекуле замещен группой ОН. Приставка "кето" также обозначает, что вместо водорода к одному из углеродов присоединен кислород (С = О), иначе ту же группу можно обозначать окончанием "он".
Биосинтез стероидов в организме осуществляется главным образом семенниками, корой надпочечников и яичниками при участии печени. Образование эстрогенов в организме происходит в основном из андрогенов (рис. 17). При этом вызывающая ароматизацию ферментная система, способная превращать андростендиол и тестостерон соответственно в эстрон и эстрадиол, распространена во многих тканях и органах (например, в жировой ткани, лимбических структурах, гипоталамусе, плаценте, волосах). Столь широкая представленность ферментной системы, обеспечивающей образование "женских" половых гормонов путем превращения андрогенов в эстрогены (наряду с выработкой последних в яичниках), создает для организма ряд преимуществ: при этом механизме гормон поступает в клетку-мишень в активном состоянии (будучи защищен от инактивирующего действия белков плазмы), и биологические ответы в клетках-мишенях получают возможность быть опосредованными внутриклеточными эстрогенами, подтверждением чему служат феномен импринтинга ферментов и рост волос на определенных участках тела.
![Рис. 17. Биосинтез андрогенов и эстрогенов [Williams R., 1974]. Пунктиром выделено образование природных эстрогенов в ткани яичников](pic/000086.jpg)
Рис. 17. Биосинтез андрогенов и эстрогенов [Williams R., 1974]. Пунктиром выделено образование природных эстрогенов в ткани яичников
Отмечена общность ранних этапов синтеза андрогенов в семенниках, яичниках и коре надпочечников. Однако, несмотря на то что в целом биосинтез стероидов одинаков во всех тканях и органах, соотношение активности разных ферментов обусловливает различные относительные количества и вид секретируемых гормонов. В интерстициальных клетках семенников, например, низка активность ферментных систем, обеспечивающих ароматизацию, и поэтому данная ткань в основном продуцирует андростендион и тестостерон. При патологии, когда процессы ароматизации усиливаются, например в случае опухолей мужских половых желез, соответственно повышаются синтез и экскреция эстрогенов [Старкова Н. Т., 1973].
В крови V. spermatica у взрослых мужчин [Hudson В. et al., 1967] были определены тестостерон, андростендион и дегидроэпи-андростерон в концентрации соответственно 47,9; 2,9 и 4,5 мкг/100 мл плазмы. В среднем яички секретируют тестостерон около 6,9 мг/сут, а кора надпочечников выделяет в больших количествах Дигидроэпиандростерон и андростендион. В тканях органов-мишеней концентрация этих гормонов выше, чем в плазме крови (так, тестостерон в больших количествах накапливается в предстательной железе и семенных пузырьках). Накопление стероидных гормонов 'в клетках-мишенях не означает, что существует какой-то особый механизм переноса их к этим клеткам: достаточно, чтобы молекулы задерживались в них. В остальных клетках тела гормон проходит через плазматическую мембрану в обоих направлениях, и его концентрация в клетке поэтому оказывается не выше, чем в крови. В тканях органов-мишеней молекулы гормона продолжают диффундировать в клетки, но лишь немногие из них выходят обратно, так что внутриклеточная концентрация их возрастает.
Секреция тестостерона у мужчин резко возрастает в период полового созревания (пубертатный период развития) и поддерживается на среднем уровне вплоть до глубокой старости. Наряду с этим, однако, даже у здоровых людей наблюдаются циклические колебания в интенсивности секреции тестостерона с периодом от 8 до 30 дней и амплитудой от 14 до 42% [Doering Ch., 1975]. Основным путем транспорта половых гормонов является кровь, хотя лимфе тоже принадлежит существенная роль в этом процессе, особенно в распространении гормонов внутри самих половых желез.
Из "мужских" половых гормонов (андрогенов) наиболее активен тестостерон. Другие натуральные андрогены (андростендион, андростерон) в 6-10 раз менее активны, чем тестостерон, а дегидроэпиандростерон и эпитестостерон - в 25-50 раз.
Мехаизм действия стероидных гормонов связан с регуляцией процессов биосинтеза белков-ферментов на уровне генов. Так, например, тестостерон, поступающий из крови в клетки, связывается специфическими белками-рецепторами, подвергается ферментной трансформации с образованием активной формы - дигидротесто-стерона, стимулирующего процесс репликации ДНК, передачи генетической информации с ДНК на РНК и биосинтез белков на рибосомах. Это вызывает усиление метаболизма в клетках органов- мишеней и в конечном счете проявляется анаболическим эффектом.
Период циркуляции половых гормонов в крови невелик, исчезновение их из крови проходит два этапа. Полупериод циркуляции в крови на первом этапе составляет 5-20 мин, а затем увеличивается до 2,5-3 ч. Выходу гормонов из крови в значительной степени способствует их поглощение тканями, где происходит их интенсивный метаболизм. Так, значительное количество стероидных гормонов диффундирует из крови в жировую ткань: она служит своего рода депо для половых гормонов, особенно для прогестерона.
Тестостерон подвергается в организме инактивации путем окисления группы ОН, связанной с 17-м атомом углерода, или путем редукции кетоновой группы, связанной с углеродом-3, до гидроксильной группы. При этом исчезает также двойная связь в кольце А (см. рис. 15). Тестостерон, образовавшийся в семенниках, превращается в малоактивные или совершенно неактивные стероидные соединения группы 17-кетостероидов (17-КС), которые выводятся из организма с мочой. Главными метаболитами тестикулярного тестостерона являются этиохоланолон, андростерон и эпиандростерон. Тестостерон, образовавшийся в коре надпочечников, превращается в дегидроэпиандростерон (рис. 18). Метаболиты тестикулярного происхождения (фракция α) составляют около 1/3, надпочечникового (фракция β) - около 2/3 общего количества 17-КС, содержащихся в моче. Метаболизм тестостерона во многом зависит от функции печени. При циррозе печени андрогенные препараты, производные тестостерона, принимаемые внутрь, полностью не инактивируются, а преобразуются в эстрогены. Эндогенный тестостерон в подобных случаях также гораздо легче превращается в эстрогены, что обусловливает развитие гинекомастии у больных с недостаточностью печени или у лиц, истощенных длительным заболеванием.
![Рис. 18. Катаболизм тестостерона в организме. I - превращение в неактивные соединения, выделяемые с мочой как 17-КС (дегидроэпиандростерон - из тестостерона надпочечникового происхождения, этиохоланолон, андростерон и эпиандростерон - из тестостерона, образовавшегося в семенниках); II - инактивация тестостерона путем синтеза эстрогенов [Teter Е., 1968]](pic/000087.jpg)
Рис. 18. Катаболизм тестостерона в организме. I - превращение в неактивные соединения, выделяемые с мочой как 17-КС (дегидроэпиандростерон - из тестостерона надпочечникового происхождения, этиохоланолон, андростерон и эпиандростерон - из тестостерона, образовавшегося в семенниках); II - инактивация тестостерона путем синтеза эстрогенов [Teter Е., 1968]
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://sohmet.ru/ 'Sohmet.ru: Библиотека по медицине'