ЭЛЕКТРОКАРДИОГРАФИЯ
Знаменитное открытие Гальвани послужило толчком к развитию учения об электричестве и всей современной электротехнике. Сам автор, однако, думал, что им открыто так называемое животное электричество. Впоследствии работы Вольта показали, что в лягушечьей лапке Гальвани имел лишь чувствительный аппарат, позволивший ему обнаружить очень слабые токи, образующиеся от соприкосновения двух разнородных проводников.
Появление электрического тока, образующегося в результате химизма жизненных процессов, было обнаружено гораздо позднее; изучение же этих явлений стало возможно только в последние десятилетия со времени устройства особо чувствительных и точных гальванометров. По существу явление, открытое Гальвани, и возникновение разницы электрических потенциалов в животных тканях очень близки, так как в основе этих процессов лежат вполне аналогичные химические или, вернее, физико-химические изменения — электролитическая диссоциация вещества.
В наиболее простой форме явление можно исследовать в системе так называемого гальванического элемента. Если мы в раствор какого-либо электролита погрузим металлическую, скажем, цинковую пластинку, то начнется растворение цинка в жидкости с образованием цинковой соли. Ионы цинка будут переходить в жидкость, унося с собою положительный заряд. Пластинка потеряет некоторое количество положительного электричества, раствор приобретает равный по величине положительный заряд. Если теперь соединить проводником пластинку с раствором, замкнуть внешнюю цепь, то в ней потечет ток от раствора к пластинке. Теоретически течение тока прекратится лишь тогда, когда будет израсходован весь запас металла, пока весь цинк не растворится.
Несколько сложнее обстоит дело при соприкосновении двух проводников второго рода, т. е. электролитов — растворов каких-либо солей или кислот. В этом случае наибольшее значение приобретают процессы диффузии. Вещества диффундируют не с одинаковой скоростью, и скорость диффузии зависит от величины частиц: наименьшие по величине частицы обладают наибольшей скоростью проникновения. Самой маленькой по величине частицей в растворе является водородный ион, и скорость его распространения наибольшая. Поэтому, если привести в соприкосновение раствор какой-либо кислоты и нейтральной соли или просто растворы одной и той же кислоты, но разных концентраций, то Н-ионы будут стремиться первыми перейти из одного более концентрированного раствора в другой, менее концентрированный. Каждый из этих растворов был электрически нейтрален, так как содержал равные между собою количества положительно и отрицательно заряженных ионов. Когда растворы приведены в соприкосновение, то, в силу законов диффузии, они будут стремиться выравнять разницу в концентрации, при чем ионы и недиссоциированные молекулы одного раствора будут проникать в другой. Но скорость проникновения различных продуктов диссоциации различна, и, в первую очередь, в раствор более низкой концентрации проникнут Н-ионы, перенеся сюда и избыток положительных зарядов. Вначале, следовательно, должна создаться некоторая разница потенциалов: более концентрированный раствор потеряет некоторое количество положительного электричества, более слабый раствор приобретет равный положительный заряд. Если замкнуть внешнюю цепь, в ней потечет электрический ток в направлении от менее концентрированного раствора к более крепкому. Сила электрического тока будет, как показал Нернст, при прочих равных условиях пропорциональна разнице концентраций растворов. На этом принципе основан способ электрометрического измерения величины кислотности жидкости путем определения концентрации свободных водородных ионов и растворе.
В результате разнообразных процессов внутриклеточного обмена в клетке образуется ряд продуктов распада кислотного характера. Наибольшее количество таких продуктов обмена образуется при мышечной работе. Из работающего участка мышцы они удаляются не сразу и на некоторой очень короткий промежуток времени создают некоторую разницу в концентрации электролитов и, следовательно, разницу электрического потенциала. Работами некоторых русских авторов установлена пропорциональность между силой электрического тока и количеством кислых продуктов обмена. Это дает основание думать, что причины появления электрического тока в работающем органе кроются в изменении химического состава клеточной протоплазмы. Схема концентрационных цепей очень удобна при изучении электрических процессов в живых тканях, но может быть справедлива для объяснения причин возникновения токов действия лишь до известной степени. Химизм тканей и процессы внутритканевого обмена еще очень мало изучены, и, надо полагать, что причины возникновения разницы электрических потенциалов гораздо сложнее.
Работающая часть мышечного волокна по отношению к части его, находящейся в состоянии покоя, приобретает отрицательный заряд. С точки зрения схемы концентрационных цепей это понятно, так как в работающей части накопляется избыток кислых продуктов, водородные ионы быстро уходят из сфгры своего возникновения, перенося положительный заряд к покоющейся части, а на месте остается избыток радикалов кислоты, имеющих отрицательный заряд. Доказать появление разницы потенциалов в работающей мышце легко, соединив ее концы с достаточно чувствительным гальванометром. Токи действия можно обнаружить во всех мышцах как поперечнополосатых, так и гладких и в сердечной мышце. Их существование доказано также для нервных клеток и волокон и для железистого аппарата. Таким образом, появление тока в мышце не должно стоять в зависимости от ее механической работы и не служит выражением последней. Электрический ток появляется в результате химических процессов в протоплазме, которые начинаются с момента появления и распространения возбуждения. И действительно, появление импульса имеет гораздо большее значение в процессе возникновения разницы электрического потенциала, чем само сокращение мышечного волокна. Экспериментально удается показать независимость электрических и механических явлений в мышце. Особенно это легко делюнстрировать на сердечной мышце. С остановленного некоторыми ядами сердца можно еще продолжительное время получать хорошо сформированную электрокардиограмму. Однако в обычных условиях распространение возбуждения и сокращение идут паралелыю, поэтому нарушения в механической работе сердца отражаются на форме ЭКГ, или, вернее, электрокардиограмма является отражением механических явлений в сердце.
Выше мы разбирали физиологические особенности сердечной мышцы. Главной особенностью этого мышечного аппарата является существование особой проводящей автономной системы. Импульс раздражения возникает и распространяется в мускулатуре сердца в строгой последовательности от одного отдела к другому. Распространение импульса требует относительно довольно продолжительного времени, и в процессе его распространения можно отметить два основных момента: распространение его в предсердной части сердечной мускулатуры и распространение его в желудочковой части. Импульс сразу охватывает всю стенку предсердий, желудочки находятся в это время в состоянии полного покоя. Р.сли исследовать в этот момент величину электрического заряда в различных отделах сердца, то окажется, как этого и можно было ожидать, исходя из теоретических предположений, что все основание сердца будет обладать отрицательным зарядом, верхушка или вообще остальная часть сердца — положительным.
Возбуждение распространяется но проводящей системе пучка Гиса, сначала достигает верхушки и трупы папиллярных мышц левого желудочка и затем передается мускулатуре основания обоих желудочков. Величина предсердий по сравнению с массой основания желудочков очень незначительна, почему практически можно принять, что в момент распространения импульса на область основания желудочков все основание сердца снова получает большой отрицательный заряд. На кривой это выражается большим острым зубцом, начинающим желудочковую часть электрокардиограммы (см. рис. 73). Затем следует небольшой период покоя и снова покатый подъем кривой, соответствующий моменту окончания систолы желудочков. В дальнейшем необходимо будет обратиться к описанию распространения электрического потенциала в сердечной мускулатуре в связи с ходом раздражения и рассмотреть более подробно связь отдельных моментов ЭКГ кривой с работой различных отделов сердечной мускулатуры.
Для более ясного понимания связи между ходом возбуждения и изменениями электрического потенциала необходимо обратиться к некоторым экспериментальным данным. Из многочисленных исследований этого рода особенного внимания заслуживают опыты Гартен-Клемента с так называемыми дифференциальными электродами. Орган или животная ткань под влиянием протекающих в ней жизненных процессов становится источником электрического тока. Наша задача заключается в исследовании этих токов. Для этого необходимо соединить участки органа с гальванометром, т. е. отвести ток в аппарат, дающий возможность судить о силе, величине и направлении электрического тока.
Производя опыты отведения тока с поверхности сердца при посредстве электродов, соприкасавшихся с сердечной мышцей на очень ограниченном пространстве, Гартен показал, что форма, направление и величина волн электрограммы стоит в тесной зависимости от направления в распространении раздражения по отношению к месту отведения. Если в каких-либо точках будут приложены отводящие ток электроды kk (см. рис. 71), возбуждение же будет распространяться в направлении ab, то электрическая кривая будет иметь форму I. Если ход раздражения будет диаметрально противоположен первому, т. е. возбуждение будет распространяться в направлении Ьа, то кривая будет иметь форму П, представляющую зеркальное изображение первого случая. При ходе возбуждения в направлении перпендикулярном линии отведения тока, т. е. в направлении cd, гальванометр не покажет никакого отклонения. При направлении возбуждения под углом к линии отведения степень отклонения гальванометра будет тем больше, чем меньше угол между линией отведения ab и линией направления возбуждения.
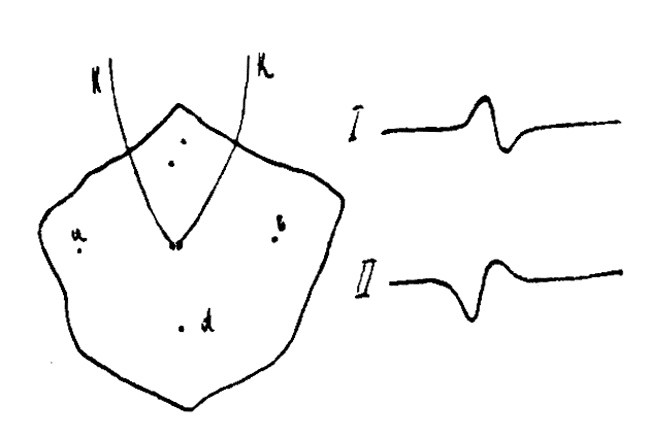
Рис. 71
При отведении токов действия у человека мы отводим их от конечностей и,таким образом,получаем не изолированное, а суммарное выражение изменений электрического потенциала всей сердечной мышцы. Напомним, что с гальванометром соединяются конечности, что может быть сделано в любом порядке. Обычно пользуются следующими комбинациями при отведении тока у человека: правая рука,леваярука — первое отведение (I); правая рука, левая нога — второе отведение (II); левая рука, левая нога — третье отведение (III).
Положение сердца и направление сердечной сси по отношению к конечностям тела несимметрично и различно в зависимости от отведения, которым приходится пользоваться. Форма и величина зубцов ЭКГ изменяется у одного и того же субъектав зависимости от отведения, в котором снята кривая. Сточки зрения опытов Гартена, это вполне понятно. Для теоретического выяснения возможных взаимоотношений между формой и направлением зубцов нормальной ЭКГ и положением сердца обратимся к схеме равнобедренного треугольника, предложенного Эйнтховеном. Для упрощения человеческое тело можно уподобить треугольнику ABC (см. рис.72 а). Углы его А и В соответствуют правой и левой верхним конечностям, С — ногам. Линия PQ представляет направление и положение сси сердца, с которой практически совпадает ход распространения возбуждения. Величина электрического потенциала, возникающего между точками РQ,будетпропорциональна проекции этой линии на сторонах треугольника и в первом отведении будет пропорциональна линии P1Q2 во втором(АС) — P2Q2,B третьем (ВС) — линии P3Q3 - Из чертежа видно, что линия P2Q2 равна сумме Р1Q1 и P3Q3. В действительности таки получается, что легко доказать экспериментально. Если сделать треугольник из какого-либо проводящего ток материала, например, из глины, и в центре его по какому-либо произвольному направлению приложить в точках PQ некоторую разность потенциалов, то в таком случае на его углах также возникнет некоторый электрический потенциал, и если мы его измерим последовательно между точками АВ, АС и ВС, то по величине потенциал между точками АС будет равен сумме потенциалов между точками между АВ и ВС.
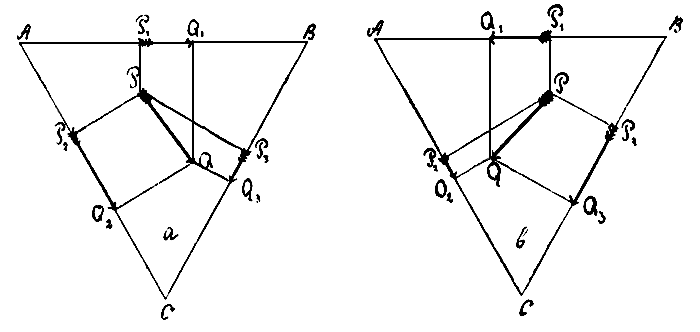
Рис. 72
Эта схема дает нам возможность объяснить разницу в величине зубцов ЭКГ, получаемой в различных отведениях. Если при одновременной съемке ЭКГ в трех отведениях все три гальванометра установлены точно на одинаковой степени чувствительности, то, измерив величину зубцов ЭКГ, можно вычислить угол наклона сердечной оси. Произведенные в этом направлении исследования показывают, что такой способ вычисления направления сердечной оси отличается большей точностью, чем ортокардиография.
Опыты Гартена показали, что не только величина, но и направление зубцов ЭКГ зависит от взаимоотношений между ходом раздражения по отношению к точкам отведения тока. Обращаясь к схеме равнобедренного треугольника, представим, что направление сердечной оси будет PQ (рис. 72 b). В таком случае при проектировании ее на стороны треугольника на стороне АВ мы будем иметь отрицательную проекцию, линия PQ будет спроектирована в направлении Q1P1, на стороне АС и ВС в положительном направлении. Подобные взаимоотношения встречаем при situs viscmim inversum и декстрокардии. В первом отведении при этом обычно получается ЭКГ с обратным направлением всех зубцов, представляющая как бы зеркальное изображение нормальной кривой; во втором и третьем отведениях положение зубцов обычно нормально (Новейшня исследованиям. Мандельштама показали, что вопрос о форме и направлении зубов ЭКГ зависит не только от направления сердечной оси, но и от изменения в расположении отделов сердца. Мандельштам различает 4 формы: I — situs viserum totalis; II - изолированная декстрокардия с зеркальным расположением сердечных полостей; III — изолированная декстрокардия с нормальным расположением полостей и сосудов; IV - смещение сердца вправо вследствни- заболевания близлежащих органов. Первые два тина вполне укладыиаютсн к схему Rinthoven'a, вторые дают ряд уклонений).
Возбуждение в сердце следует от основания к верхушке. В этом же направлении следуют главнейшие изменения в распределении электрического потенциала. При снимании ЭКГ мы можем гальванометр включить в цепь в любом паправлении, но условно принято его включать так, чтобы в момент появления отрицательного потенциала у основания сердца при регистрации отклонений гальванометра зубцы кривой были направлены вверх и чтение ее производилось слева направо.
При этих условиях появление ипмульса в синусе и время его распространения по мускулатуре предсердий характеризуется появлением небольшого, направленного вверх положительного зубца, носящего по номенклатуре Эйнтховена название зубца Р (см. рис. 73). Величина его в нормальных условиях соответствует разнице электрического потенциала примерно в 0,1—0,2 милливольта. При отведении с конечностей лучше всего Р различимо во втором отведении, что понятно из вышеизложенного. Длительность Р периода равна приблизительно 0,1 секунды. Вершина его заострена, и иногда можно отметить небольшое ее раздвоение, что, вероятно, стоит в связи с неодновременным распространением импульса по правому и левому предсердию. При некоторых патологических условиях Р может быть отрицательным, что объясняется ретроградным течением импульса от предсердно-желудочковой перегородки к синусу. Действительно, в этом случае в первый момент максимум отрицательного потенциала должен смещаться по направлению к верхушке сердца и, исходя из схемы треугольника, проекция направления хода возбуждения на сторонах его в этом случае будет обратная. Таковой она обычно остается для всех трех отведений, однако при почти горизонтальном направлении сердечной оси Р теоретически должно быть в первом и втором отведениях отрицательным, а в третьем положительным. При патологических условиях в ходе возбуждения действительно приходится сталкиваться с различным направлением Р.
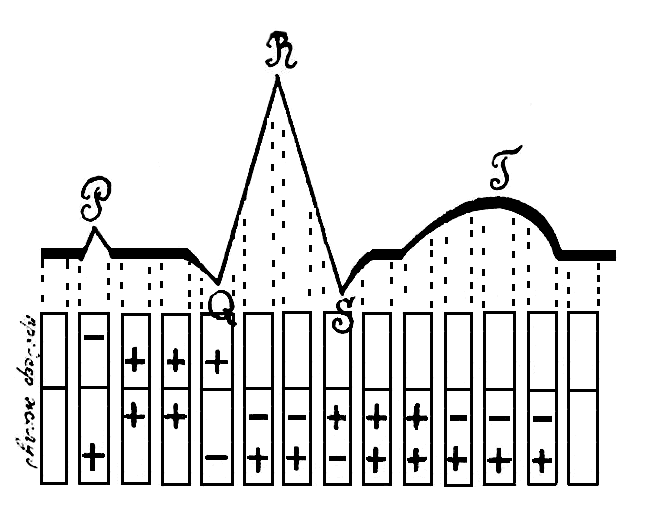
Рис. 73. Сопоставление элекрокардиографическон кривой с изменениями электрического потенциала сердечной мышцы. Сверху кривая PQRST, внизу распределение потенциала в предсердиях и желудочках
Зубец Р имеет большое практическое значение, так как он показывает время появления и последовательность выработки синусальных импульсов и отмечает ход сокращения предсердий.
Вслед за зубцом Р следует короткий период электрического покоя, соответствующий времени проведения импульса в желудочковой части специфической системы. На ЭКГ это время характеризуется горизонтальным направлением кривой на протяжении между окончанием зубца Р и началом желудочкового комплекса, почти совпадающей с нулевой линией потенциала. Время электрического покоя — проведения колеблется между 0,1 и 0,17 секунды, т. е. в довольно больших границах. Величина времени проведения несколько различна в разных отведениях и наибольшая — во втором. Она колеблется у одного и того же субъекта в зависимости от частоты сердечных сокращений, уменьшаясь при тахикардии и увеличиваясь при брадикардии.
В течение времени проведения электрокардиографическая кривая принимает горизонтальное направление, однако это не означает полного электрического покоя, так как уровень кривой в этот момент никогда не достигает нулевой линии, но лежит обычно немного ниже. Горизонтальное направление кривая принимает, очевидно, потому, что разыгрывающиеся в этот промежуток времени процессы симметрично расположены по отношению к остальной массе сердца и по отношению к отводящим ток электродам.
Желудочковый комплекс ЭКГ начинается небольшим отрицательным колебанием, носящим название зубца Q. Он непосредственно переходит в большой острый положительный зубец R, который в свою очередь непосредственно переходит в небольшой отрицательный зубец S. Комплекс QRS составляет начальную группу желудочкового периода ЭКГ. Затем кривая принимает почти горизонтальное направление и плавно переходит в покатый финальный зубец Т.
Для обозначения волн электрокардиографической кривой общепринятой в настоящее время является номенклатура Эйнтховена по порядку последних букв алфавита: Р, Q, R, S, Т. Другая номенклатура, предложенная F. Kraus H'Nicolai, также нередко встречается, особенно в немецкой литературе. Предсердный зубец обозначается буквой А, зубец R носит название начального (Jnitialschwankung) и обозначается буквой I, зубец Т — конечного колебания и обозначается буквой F (Finalschwaukung). Второстепенные колебания Q и S обозначаются соответственно a J(i) и (i)F.
Относительно промежутка Р — Q было уже упомянуто, что горизонтальное направление кривой в этом случае не говорит за электрический покой. Этот момент соответствует передаче импульса через самый узел Тавара. Появление отрицательного колебания Q указывает на появление отрицательного электрического потенциала у верхушки сердца (рис. 73). Это стоит, видимо, в связи с распространением раздражения по специфической системе. По протяжению длина ветвей Гисовского пучка, считая от узла Тавара до верхушки сердца, составляет около 3 см. Если принять в расчет скорость распространения возбуждения 3 м. в секунду, то от момента перехода возбуждения от узла к ветвям до момента достижения им верхушки сердца должно пройти 0,01 секунды. Время от начала волны Q до ее верхушки как раз и равно 0,01 секунды. В дальнейшем возбуждение начинает распространяться по восходящим ветвям системы, длина которых составляет около 6-8 см., следовательно, для достижения основания желудочков раздражению потребуется около 0,027 секунды. Это время соответствует протяжению от вершины Q до вершины R.
Таким образом, между временем распространения возбуждения и временем появления начальных колебаний вентрикулярного комплекса имеется тесная зависимость. Благодаря ряду точных экспериментальных исследований,установлено, что между распространением возбуждения и началом сокращения проходит чрезвычайно малый промежуток времени, измеряемый тысячными долями секунды. Первые механические явления в желудочках начинаются сейчас же после перехода Q в восходящую часть R. В этот момент появляются начальные колебания первого тона, и отмечается небольшое повышение внутрижелудочкового давления. Максимум звуковых колебаний первого тока совпадает с нисходящей частью R. Зубец S указывает, что вслед за начальным отрицательным зарядом в период Q верхушка еще раз приобретает отрицательный потенциал (см. рис. 73). Это ставят в зависимость от распространения возбуждения в самой мускулатуре желудочков. Возбуждение вначале достигает, главным образом, мышечных волокон с сердечной верхушки, эти последние приходят первыми в наиболее интенсивное возбуждение, затем раздражение передается в течение короткого времени всей остальной массе желудочковой мускулатуры, и все сердце приобретает равномерный отрицательный потенциал. Кривая приобретает на короткое время горизонтальное направление и расположена чаще выше нулевой линии.
Постепенно мышечные волокна желудочковой стенки начинают приходить в состояние покоя. При этом процесс опять начинается с верхушки — части, ранее других приходящей в состояние возбуждения. Основание желудочков более массивно, почему дольше сохраняется в состоянии возбуждения. Снова появляется перевес отрицательного потенциала в области основания сердца по отношению к пришедшей уже в состояние покоя верхушке. На кривой это выражается появлением финального положительного зубца Т. Таким образом, волна Т выражает ход процессов в самом миокарде желудочков и, так как по сравнению со специфической системой они протекают здесь гораздо медленнее, то подъем и спадение финального колебания отличаются плавностью. Вершина Т совпадает с появлением второго тона.
По сравнению с зубцом Р, который отличается большим постоянством как но величине, так и по форме желудочковый комплекс QRT индивидуально очень различен — он дает значительные изменения в зависимости от отведения, в котором снята кривая. Но для данного отведения форма и взаимоотношения зубцов индивидуально очень характерны и сохраняют свои особенности в течение всей жизни субъекта. Изменения ЭКГ наступают только после повреждения проводящих путей или самого миокарда.
Предсердия не имеют специфической проводящей системы, и импульс в их стенке от узла К.-Ф. распространяется равномерно по радиусам во все стороны и, следовательно, одинаков в любом сердце, почему и нельзя отметить каких-либо индивидуальных особенностей в структуре зубца Р.
Проводящая система желудочковой части представляет заметные индивидуальные колебания в своей анатомической структуре, поэтому порядок распространения импульса индивидуально несколько разнится.
Выше мы разбирали колебания электрического потенциала в зависимости от места отведения электрическою тока. На ЭКГ величина разницы потенциала выражается высотой зубцов, и правило, что II = I + III, справедливо главным образом по отношению к величине R. На величину и положение Q и S большее влияние оказывает топографическое расположение ветвей проводящей системы. Эти зубцы могут быть выражены хорошо или совершенно отсутствовать у здоровых людей. Вообще эти зубцы отличаются большим непостоянством, и у одного и того же субъекта они могут быть хорошо выражены в одном отведении и отсутствовать в другом. Как правило, они лучше выражены но втором отведении.
R — обычно наиболее высокий зубец ЭКГ и по величине соответствует разнице потенциалов в 5,5 до 16,5 милливольта. Величина его не зависит, как некоторые предполагали, от массы и силы желудочковой мускулатуры. Продолжительность всего периода QR колеблется в пределах между 0,06 и 0,1 секунды. Промежуток между S и Т колеблется в пределах между 0,03 до 0,15 секунды.
Зубец Т в I и II отведении всегда положителен, в III отведении он нередко бывает отрицательным, что в значительной мере зависит от направления сердечной оси и наблюдается при горизонтальном расположении сердца. Форма его, продолжительность и направление в значительной степени зависят от состояния миокарда и его сократительных свойств. Продолжительность Т периода значительно колеблется даже при небольших изменениях скорости сердечных сокращений. При ритме 60 ударов в минуту продолжительность периода Т около 0,32 секунды, при ритме 90 ударов — 0,28 секунды.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://sohmet.ru/ 'Sohmet.ru: Библиотека по медицине'